 2015-05-26
2015-05-26 3812
3812
В этих условиях происходит восстановление NAD+, рибофлавина, хинона, Fe3+, кислорода. Эти реакции получили название «реакций Красповского». Таким образом, молекула хлорофилла может выступать не только в роли первичного акцептора электрона, но и в роли его первичного донора.
Из всего сказанного следует, что молекула хлорофилла благодаря структурным и физико-химическим особенностям способна выполнять три важнейшие функции: 1) избирательно поглощать энергию света, 2) запасать ее в виде энергии электронного возбуждения, 3) фотохимически преобразовывать энергию возбужденного состояния в химическую энергию первичных фотовосстановленных и фотоокисленных соединений.
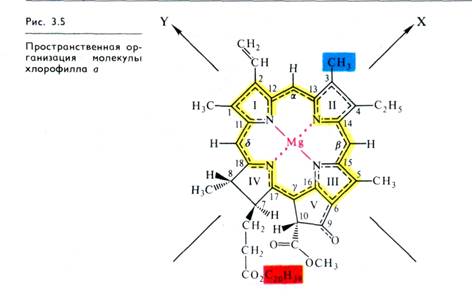 |
Для функционирования молекулы хлорофилла в процессах фотосинтеза существенное значение имеет ее пространственная организация. Магний-порфириновое кольцо молекулы представляет собой почти плоскую пластинку толщиной 0,42 нм и площадью 1 нм2. Это гидрофильная часть молекулы хлорофилла. Длинный алифатический остаток фитола (2 нм), обра зующий угол с порфириновым кольцом,— ее гидрофобный полюс (рис. 3.5), необходимый для взаимодействия молекулы хлорофилла с гидрофобными участками мембранных белков и липидов.
 Синезеленые водоросли (цианобактерии), красные морские водоросли и некоторые морские криптомонады помимо хло рофилла а и каротиноидов содержат пигменты фикобилины. Наиболее известные представители фикобилинов — фикоэритро- билины и фикоцианобилины. Первые преобладают у красных водорослей и определяют их цвет, вторые — у синезеленых.
Синезеленые водоросли (цианобактерии), красные морские водоросли и некоторые морские криптомонады помимо хло рофилла а и каротиноидов содержат пигменты фикобилины. Наиболее известные представители фикобилинов — фикоэритро- билины и фикоцианобилины. Первые преобладают у красных водорослей и определяют их цвет, вторые — у синезеленых.
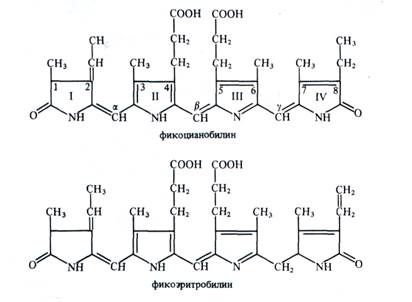
Структура и свойства фикобилинов. По структуре фикобилины (от греч. «phycos» — водоросль и лат. bilis — желчь) относятся к группе желчных пигментов — билинов (у животных представитель этой группы — билирубин). Это тетрапирролы с открытой цепью, имеющие систему конъюгированных двойных и одинарных связей. В своем составе они не содержат атомов магния или других металлов, а также фитола.
У фикоцианобилина пиррольные кольца соединены между собой метиновыми мостиками. I и IV пирролы имеют по одной карбонильной группе. Пиррольные кольца содержат следующие боковые радикалы: четыре метальных (у С1,|3,6,7), винильную (у С2), этильную (у С8) и два остатка пропионовой кислоты (у С4 и С5).
Фикобилины являются хромофорными группами фикобили-протеинов — глобулиновых белков, с которыми в отличие от хлорофиллов они связаны прочными ковалентными связями. Фикобилипротеины делятся на три основные группы: 1) фикоэритрины — белки красного цвета с максимумом поглощения от 498 до 568 нм, 2) фикоцианины — сине-голубые белки с максимумами поглощения от 585 до 630 нм, 3) аллофикоцианины — синие белки с максимумами поглощения от 585 до 650 нм. Все эти хромопротеины обладают флуоресценцией с максимумами 575 — 578, 635 — 647 и 660 нм соответственно.
Фикобилипротеины водорастворимы, в клетках водорослей они локализованы в фикобилисомах — гранулах, расположенных на наружной поверхности фотосинтетических ламелл.
Значение фикобилинов. Максимумы поглощения света у фикобилинов находятся между двумя максимумами поглощения у хлорофилла: в оранжевой, желтой и зеленой частях спектра (см. рис. 3.2). Значение такого распределения максимумов поглощения становится понятным, если вспомнить оптические свойства воды, которая поглощает прежде всего длинноволновые лучи. На глубине 34 м в морях и океанах полностью исчезают красные лучи, на глубине 177 м — желтые, на глубине 322 м — зеленые и, наконец, на глубину свыше 500 м не проникают даже синие и фиолетовые лучи. В связи с таким изменением качественного состава света в верхних слоях морей и океанов обитают преимущественно зеленые водоросли, глубже — синезеленые и еще глубже — водоросли с красной окраской. В. Т. Энгельман назвал это явление хроматической комплементарной адаптацией водорослей. По его наблюдениям (1881 — 1884), наиболее интенсивная ассимиляция СО2 у водорослей с различной окраской соответствует максимумам поглощения света пигментными системами этих водорослей. Русский исследователь Н. М. Гайдуков (1903) экспериментально показал, что если культуру синезеленой водоросли Oscillaria sancta выращивать на свету разного спектрального состава, то у нее развивается дополнительная (комплементарная) окраска. При освещении зеленым светом водоросли становятся оранжево-красными, а при действии красных лучей — зелеными.
В настоящее время известно, что эти изменения цвета клеток связаны с изменениями в синтезе фикобилинов, принимающих участие в процессе фотосинтеза. Таким образом, у водорослей фикобилины — дополнительные пигменты, выполняющие вместо хлорофилла b функции светособирающего комплекса. Около 90 % энергии света, поглощенного фикобилинами, передается на хлорофиллы а (см. 3.3.1). Явление хроматической комплементарной адаптации обнаружено далеко не у всех видов синезеленых и красных водорослей. У многих из них адаптация к изменяющемуся спектральному составу света обеспечивается изменением количества и состава хлорофиллова.
Кроме фикобилинов, участвующих в фотосинтезе у водорослей, у всех растений имеется другой фикобилин — фитохром, являющийся фоторецептором для восприятия красного и дальнего красного света и выполняющий регуляторные функции (см. 2.7.2).
 Каротиноиды — жирорастворимые пигменты желтого, оранжевого, красного цвета — присутствуют в хлоропластах всех растений. Они входят также в состав хромопластов в незеленых частях растений, например в корнеплодах моркови, от латинского наименования которой (Daucus carota L.) они и получили свое название. В зеленых листьях каротиноиды обычно незаметны из-за присутствия хлорофилла, но осенью, когда хлорофилл разрушается, именно каротиноиды придают листьям характерную желтую и оранжевую окраску. Каротиноиды синтезируются также бактериями и грибами, но не животными организмами. В настоящее время известно около 400 пигментов, относящихся к этой группе.
Каротиноиды — жирорастворимые пигменты желтого, оранжевого, красного цвета — присутствуют в хлоропластах всех растений. Они входят также в состав хромопластов в незеленых частях растений, например в корнеплодах моркови, от латинского наименования которой (Daucus carota L.) они и получили свое название. В зеленых листьях каротиноиды обычно незаметны из-за присутствия хлорофилла, но осенью, когда хлорофилл разрушается, именно каротиноиды придают листьям характерную желтую и оранжевую окраску. Каротиноиды синтезируются также бактериями и грибами, но не животными организмами. В настоящее время известно около 400 пигментов, относящихся к этой группе.
Структура и свойства. Элементарный состав каротиноидов установил Вильштеттер. С 1920 по 1930 г. была определена структура основных пигментов этой группы. Искусственный синтез ряда каротиноидов впервые осуществлен в 1950 г. в лаборатории П. Каррера. К каротиноидам относятся три группы соединений: 1) оранжевые или красные пигменты каротины (С40Н56); 2) желтые ксантофиллы (C40H56O2 и С40Н56О4); 3) каротиноидные кислоты — продукты окисления каротиноидов с укороченной цепочкой и карбоксильными группами (например, С2оН24О4 ~ кроцетин, имеющий две карбоксильные группы).
Каротины и ксантофиллы хорошо растворимы в хлороформе, бензоле, сероуглероде, ацетоне. Каротины легко растворимы в петролейном и диэтиловом эфирах, но почти нерастворимы в метаноле и этаноле. Ксантофиллы хорошо растворимы в спиртах и значительно хуже в петролейном эфире.
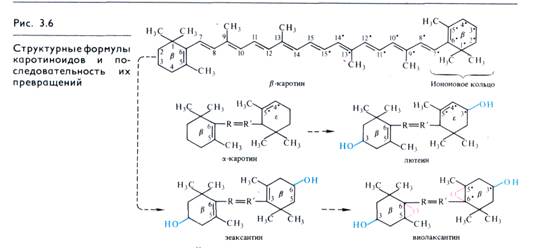
Все каротиноиды — полиеновые соединения. Каротиноиды первых двух групп состоят из восьми остатков изопрена, которые образуют цепь конъюгированных двойных связей. Каротиноиды могут быть ациклическими (алифатическими), моно- и бициклическими. Циклы на концах молекул каротиноидов являются производными ионона (рис. 3.6). Примером ациклического каротиноида может служить ликопин (С40Н56) —
основной каротин некоторых плодов (в частности, томатов) и пурпурных бактерий.
β-kаротин (рис. 3.6) имеет два β -иононовых кольца (двойная связь между С5 и С6). При гидролизе β -каротина по центральной двойной связи образуются две молекулы витамина А (ретинола). α -каротин отличается от β -каротина тем, что у него одно кольцо β -иононовое, а второе — ε-иононовое (двойная связь между С4 и С5).
Ксантофилл лютеин — производное α -каротина, а зеаксан-тик — β -каротина. Эти ксантофиллы имеют по одной гидро-ксильной группе в гаждом иононовом кольце. Дополнительное включение в молекулу зеаксантина двух атомов кислорода по двойным связям С5—С6 (эпоксидные группы)
«виолаксантин» связано с выделением этого соединения из лепестков желтых анютиных глазок (Viola tricolor). Зеаксантин впервые получен из зерновок кукурузы (Zea mays). Лютеин (от лат. luteus — желтый) содержится, в частности, в желтке куриных яиц. К наиболее окисленным изомерам лютеина относится фукоксантин (С40Н60О6) — главный ксантофилл бурых водорослей.
Основные каротиноиды пластид высших растений и водорослей — β-каротин, лютеин, виолаксантин и неоксантин. Синтез каротиноидов начинается с ацетил-СоА через мевалоновую кислоту, геранилгеранилпирофосфат до ликопина, который является предшественником всех других каротиноидов. Синтез каротиноидов происходит в темноте, но резко ускоряется при действии света. Спектры поглощения каротиноидов характеризуются двумя полосами в фиолетово- синей и синей области от 400 до 500 нм (см. рис. 3.2). Количество и положение максимумов поглощения зависят от растворителя. Этот спектр поглощения определяется системой конъюгированных двойных связей. При увеличении числа таких связей максимумы поглощения смещаются в длинноволновую область спектра.
Каротиноиды, как и хлорофиллы, нековалентно связаны с белками и липидами фотосинтетических мембран.
Роль каротиноидов в процессах фотосинтеза. Каротиноиды — обязательные компоненты пигментных систем всех фотосинтезирующих организмов. Они выполняют ряд функций, главные из которых: 1) участие в поглощении света в качестве дополнительных пигментов, 2) защита молекул хлорофиллов от необратимого фотоокисления. Возможно, Каротиноиды принимают участие в кислородном обмене при фотосинтезе.
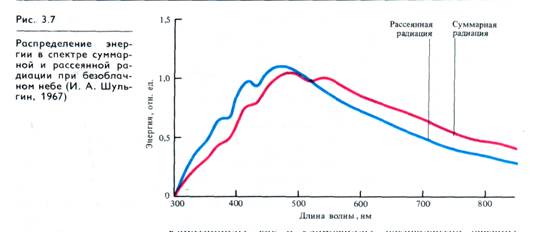
Важное значение каротиноидов как дополнительных пигментов, поглощающих свет в сине-фиолетовой и синей частях спектра, становится очевидным при рассмотрении распределения энергии в спектре суммарной солнечной радиации на поверхности Земли. Как следует из рис. 3.7, максимум этой радиации приходится на сине-голубую и зеленую части спектра (480 — 530 нм). В естественных условиях доходящая до поверхности Земли суммарная радиация слагается из потока прямой солнечной радиации на горизонтальную поверхность и рассеянной радиации неба. Рассеивание света в атмосфере происходит благодаря аэрозольным частицам (капли воды, пылинки и т. д.) и флуктуациям плотности воздуха (молекулярное рассеяние). Спектральный состав суммарной радиации в области 350 — 800 нм при безоблачном небе в течение дня почти не меняется. Объясняется это тем, что увеличение доли красных лучей в прямой солнечной радиации при низком стоянии Солнца (см. рис. 3.25) сопровождается увеличением доли рассеянного света, в котором много сине-фиолетовых лучей. Атмосфера Земли в значительно большей степени рассеивает лучи коротковолновой части спектра (интенсивность рассеяния обратно пропорциональна длине волны в четвертой степени), поэтому небо выглядит голубым. При отсутствии прямого солнечного света (пасмурная погода) увеличивается доля сине-фиолетовых лучей. Эти данные указывают на важность коротковолновой части спектра при использовании наземными растениями рассеянного света и возможность участия каротиноидов в фотосинтезе в качестве дополнительных пигментов. В модельных опытах показана высокая эффективность переноса энергии света от каротиноидов к хлорофиллу а, причем этой способностью обладают молекулы каротинов, но не ксантофиллов.
 Вторая функция каротиноидов — защитная. Впервые данные о том, что каротиноиды могут защищать молекулы хлорофилла от разрушения, были получены Д. И. Ивановским (1913). В его опытах пробирки, содержащие одинаковый объем раствора хлорофилла и разные концентрации каротиноидов, выставлялись на 3 ч на прямой солнечный свет. Оказалось, что чем больше каротиноидов было в пробирке, тем в меньшей степени разрушался хлорофилл. В дальнейшем эти данные получили многочисленные подтверждения. Так, бескаротиноидные мутанты хламидомонады на свету в атмосфере кислорода погибают, а в темноте при гетеротрофном способе питания нормально развиваются и размножаются. У мутанта кукурузы, у которого отсутствовал синтез каротиноидов, образующийся хлорофилл в аэробных условиях при сильном освещении быстро разрушался. В отсутствие кислорода хлорофилл не разрушался.
Вторая функция каротиноидов — защитная. Впервые данные о том, что каротиноиды могут защищать молекулы хлорофилла от разрушения, были получены Д. И. Ивановским (1913). В его опытах пробирки, содержащие одинаковый объем раствора хлорофилла и разные концентрации каротиноидов, выставлялись на 3 ч на прямой солнечный свет. Оказалось, что чем больше каротиноидов было в пробирке, тем в меньшей степени разрушался хлорофилл. В дальнейшем эти данные получили многочисленные подтверждения. Так, бескаротиноидные мутанты хламидомонады на свету в атмосфере кислорода погибают, а в темноте при гетеротрофном способе питания нормально развиваются и размножаются. У мутанта кукурузы, у которого отсутствовал синтез каротиноидов, образующийся хлорофилл в аэробных условиях при сильном освещении быстро разрушался. В отсутствие кислорода хлорофилл не разрушался.
Каким же образом каротиноиды препятствуют разрушению хлорофилла? В настоящее время показано, что каротиноиды способны реагировать с хлорофиллом, находящимся в три-плетном состоянии, предотвращая его необратимое окисление. При этом энергия триплетного возбужденного состояния хлорофилла превращается в теплоту:
ХлТ*+ Каротиноид——> ХлS0 + КаротиноидТ* КаротиноидТ*——> КаротиноидS0 + Теплота
Кроме этого каротиноиды, взаимодействуя с возбужденным (синглетным) кислородом, который неспецифически окисляет многие органические вещества, могут переводить его в основное состояние:
Каротиноид + 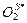 ——> КаротиноидТ* + О2 Каротиноид Т* —> Каротиноид S0 + Теплота
——> КаротиноидТ* + О2 Каротиноид Т* —> Каротиноид S0 + Теплота
Менее ясна роль каротиноидов в кислородном обмене при фотосинтезе. У высших растений, мхов, зеленых и бурых водорослей осуществляется светозависимое обратимое дез-эпоксидирование ксантофиллов. Примером такого превращения может служить виолаксантиновый цикл:
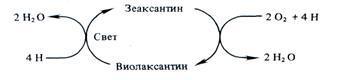
Значение виолаксантинового цикла остается невыясненным. Возможно, он служит для устранения излишков кислорода. Каротиноиды у растений выполняют и другие функции, не связанные с фотосинтезом. В светочувствительных «глазках» одноклеточных жгутиковых и в верхушках побегов высших растений каротиноиды, контрастируя свет, способствуют определению его направления. Это необходимо для фототаксисов у жгутиковых и фототропизмов у высших растений.
Каротиноиды обусловливают цвет лепестков и плодов у некоторых растений1. Производные каротиноидов — витамин А, ксантоксин, действующий подобно АБК, и другие биологически активные соединения. Хромопротеин родопсин, обнаруженный у некоторых галофильных бактерий, поглощая свет, функционирует в качестве Н+-помпы. Хромофорной группой бактериородопсина является ретиналь — альдегидная форма витамина А. Бактериородопсин аналогичен родопсину зрительных анализаторов животных.
 На световой фазе фотосинтеза происходит поглощение света молекулами хлорофилла а c участием дополнительных пигментов (хлорофилла b, каротиноидов, фикобилинов) и трансформация энергии света в химическую энергию АТР и восстановленного NADPH. Все эти процессы осуществляются в фотохимически активных мембранах хлоропластов (см. 1.1.2) и представляют собой сложную систему фотофизических, фотохимических и химических реакций, природа которых в настоящее время в значительной степени расшифрована.
На световой фазе фотосинтеза происходит поглощение света молекулами хлорофилла а c участием дополнительных пигментов (хлорофилла b, каротиноидов, фикобилинов) и трансформация энергии света в химическую энергию АТР и восстановленного NADPH. Все эти процессы осуществляются в фотохимически активных мембранах хлоропластов (см. 1.1.2) и представляют собой сложную систему фотофизических, фотохимических и химических реакций, природа которых в настоящее время в значительной степени расшифрована.
В состав ламелл хлоропластов входят пять многокомпонентных белковых комплексов: светособирающий комплекс, фотосистемы I и II, цитохромный комплекс, включающий цитохромы b6 и ƒ, и АТРазный комплекс, участвующий в синтезе АТР. Все они функционируют, взаимодействуя друг с другом.
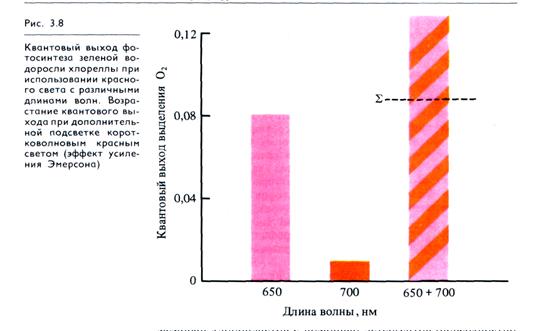
 Две фотосистемы. Впервые идею о существовании в хлоро-пластах двух фотосистем высказал Р. Эмерсон (1957), изучая влияние света на квантовый выход фотосинтеза у хлореллы. Под квантовым выходом фотосинтеза понимается количество выделившегося О2 или связанного СО2 на 1 квант поглощенной энергии. Было показано, что квантовый выход высок при освещении хлореллы красными лучами с длиной волны 660-680 нм. Использование красного света с большей длиной волны приводило к снижению квантового выхода, а при 700 нм фотосинтез почти прекращался, хотя эта часть спектра еще поглощается хлорофиллом. Однако если хлореллу одновременно освещали коротковолновым (650 нм) и длинноволновым (700 нм) красным светом, то суммарный эффект (Σ) был выше, чем при действии каждого красного света в отдельности (рис. 3.8). Это явление получило название эффекта усиления Эмерсона. Отсюда возникло предположение, что в хлоропластах взаимодействуют две пигментные системы.
Две фотосистемы. Впервые идею о существовании в хлоро-пластах двух фотосистем высказал Р. Эмерсон (1957), изучая влияние света на квантовый выход фотосинтеза у хлореллы. Под квантовым выходом фотосинтеза понимается количество выделившегося О2 или связанного СО2 на 1 квант поглощенной энергии. Было показано, что квантовый выход высок при освещении хлореллы красными лучами с длиной волны 660-680 нм. Использование красного света с большей длиной волны приводило к снижению квантового выхода, а при 700 нм фотосинтез почти прекращался, хотя эта часть спектра еще поглощается хлорофиллом. Однако если хлореллу одновременно освещали коротковолновым (650 нм) и длинноволновым (700 нм) красным светом, то суммарный эффект (Σ) был выше, чем при действии каждого красного света в отдельности (рис. 3.8). Это явление получило название эффекта усиления Эмерсона. Отсюда возникло предположение, что в хлоропластах взаимодействуют две пигментные системы.
Предположение Эмерсона о двух пигментных системах подтвердилось в последующих работах, в которых из мембран хлоропластов с помощью детергентов (поверхностно-активных веществ, диссоциирующих гидрофобные связи), дифференциального центрифугирования в градиенте плотности сахарозы и других приемов удалось выделить и изучить белковые комплексы фотосистемы I (ФС I) и фотосистемы II (ФС II).
1 Однако в большинстве случаев цвет лепестков зависит от присутствия антоцианов, локализованных в вакуолях.
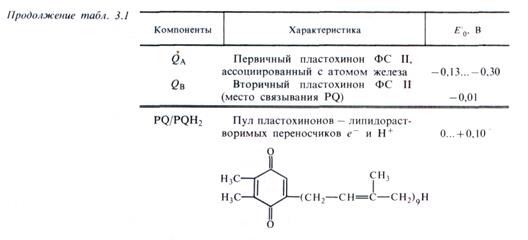
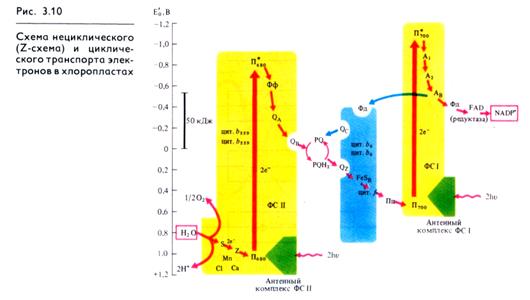
В состав ФС I в качестве реакционного центра входит димер пигмента П700 (хлорофилл а с максимумом поглощения 700 нм), а также хлорофиллы а675-695, играющие роль антенного компонента фотосистемы I (табл. 3.1, см. рис. 3.10). Первичным акцептором электронов в этой системе является мономерная форма хлорофилла а695 (A1), вторичными акцепторами — А2 и Ав (железосерные белки — FeS). Комплекс ФС I под действием света восстанавливает водорастворимый FeS-белок ферредоксин (Фд) и окисляет медьсодержащий водорастворимый белок пластоцианин (Пц).
Белковый комплекс ФС II включает в себя реакционный центр, содержащий хлорофилл а П680, а также антенные пигменты — хлорофиллы а 670-683. Первичным акцептором электронов в этой фотосистеме выступает феофетин а (Фф), передающий электроны на первичный пластохинон, ассоциированный с FeS (QA) и дальше на вторичный пластохинон (QB). В состав ФС II входят также белковый комплекс S-системы, окисляющий воду, и переносчик электронов Z, связанный с системой S и служащий донором е- для П680. Этот комплекс функционирует с участием марганца, хлора и кальция. В ФС II локализован цитохром b559. ФС II восстанавливает пластохинон (PQ) и окисляет воду с выделением О2 и протонов.
Связующим звеном между ФС II и ФС I служат пул (фонд) пластохинонов, белковый цитохромный комплекс b6 — f и пластоцианин.
В ходе эволюции ФС I возникла раньше. Она функционирует у ныне существующих фотосинтезирующих бактерий, фотосинтез которых осуществляется без разложения воды и выделения О2 (фоторедукция). В этом случае функцию доноров электронов для световой фазы фотосинтеза выполняют легко окисляемые соединения, такие, как H2S, H2, СН4 и др.
Антенные комплексы и миграция энергии в пигментных системах. Эмерсон и Арнольд показали, что в хлоропластах растений на каждый реакционный центр приходится от 200 до 400 молекул хлорофилла, функции которых связаны с поглощением света и передачей энергии возбужденного состояния на реакционные центры. Молекулы хлорофилла а и дополнительные пигменты — хлорофилл b, каротиноиды, фико-билины — входят в состав антенных, или светособирающих, комплексов (ССК). Из ламелл хлоропластов выделены свето-собирающий белковый комплекс с хлорофиллами а и b (ССК a-b), тесно связанный с ФС II, и антенные комплексы, непосредственно входящие в фотосистемы I и II (фокусирующие антенные компоненты фотосистем).
В ССК a-b присутствуют хлорофилл а в двух или трех формах с максимумами поглощения между 660 и 675 нм, хлорофилл b с максимумом поглощения 650 нм и каротиноиды. В каждом ССКа_ь содержится от 120 до 240 молекул хлорофиллов, причем отношение хлорофилла а к хлорофиллу b составляет 1,2—1,4. Половина белка тилакоидов и около 60% общего количества хлорофилла локализовано в ССК. У сине-зеленых и красных водорослей, у которых хлорофилл b отсутствует, роль ССК выполняют фикобилисомы, в состав которых входят фикобилины.

Антенный белковый комплекс ФС II содержит 40 молекул хлорофиллов а с максимумами поглощения 670 — 683 нм на один П680 и β-каротин.
Антенный белковый комплекс ФС I состоит из хромопротеи-нов, содержащих 110 молекул хлорофиллов а с максимумами поглощения 680 — 695 нм на один П700 из них 60 молекул — компоненты антенного комплекса самой фотосистемы, а 50 входят в состав комплекса, который можно рассматривать как ССК ФС I. Антенный комплекс ФС I также содержит Р-каротин.
Хромопротеины антенных комплексов не обладают фотохимической и энзиматической активностью. Если бы каждая молекула хлорофилла преобразовывала поглощенную ею энергию кванта света в фотохимическую реакцию, то такая система была бы крайне нерентабельна. Продолжительность синглетного возбужденного состояния исчисляется 10-12 — 10-9 с, и даже на прямом солнечном свету 1 квант света поглощается молекулой хлорофилла не чаще одного раза за 0,1 с. Большую часть времени молекула хлорофилла «простаивает». Поэтому роль пигментов антенных комплексов состоит в том, чтобы собирать и передавать энергию квантов на небольшое количество молекул реакционных центров П680 и П700, которые и осуществляют фотохимические реакции. Аналогично отдельные капли дождя ударяют в крышу и, сливаясь, создают постоянный ток воды в водостоке.
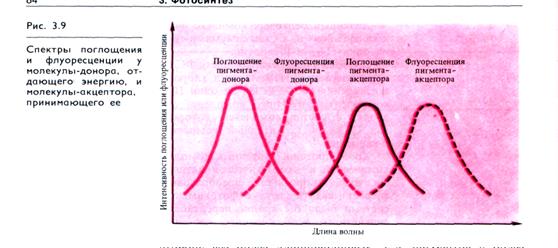
Передача (миграция) энергии по пигментам антенных комплексов происходит по принципу индуктивного резонанса (без флуоресценции и переноса заряда). Природа индуктивного резонанса состоит в следующем. Каждая молекула хлорофилла, поглотившая квант света и перешедшая в синглетное возбужденное состояние, является молекулярным осциллятором. Возникающее вокруг возбужденной молекулы переменное электрическое поле с определенной частотой колебаний индуцирует осцилляцию диполя (электрон — ядро) соседней молекулы. При этом молекула-донор переходит в основное состояние, а молекула-акцептор — в возбужденное. Механической моделью такой резонансной передачи энергии может быть система из двух маятников, связанных слабой пружинкой. Раскачивание одного из маятников вызывает колебания другого и затухание колебаний первого.
Условиями для резонансного переноса энергии электронного возбуждения служат малые расстояния между молекулами, не превышающие 10 нм, и перекрытие частот колебаний у двух взаимодействующих молекул, о чем можно судить по степени наложения спектра флуоресценции возбужденной донорной молекулы и спектра поглощения молекулы-акцептора (рис. 3.9). Флуоресценция каждой молекулы-донора имеет более длинноволновый максимум по сравнению с максимумом ее поглощения (в соответствии с правилом Стокса — см. 3.2.1) и в большей или меньшей степени перекрывает зону поглощения акцепторной молекулы. Миграция энергии осуществляется от коротковолновых пигментов в сторону все более длинноволновых, т. е. пигментов с более низким уровнем синглетного возбужденного состояния.
В антенных комплексах перенос энергии осуществляется в ряду: каротин (400 — 550 нм)——> хлорофилл b (650 нм)——>хлорофиллы а (660 — 675 нм)——> П680 (ФС II). Скорость резонансного переноса энергии от молекулы к молекуле 10-10 — 10-9 с, причем эффективность переноса между молекулами хлорофилла достигает 100, а между молекулами каротина и хлорофилла — лишь 40%. У синезеленых и красных водорослей резонансная передача энергии происходит в следующем порядке: фикоэритрин (570 нм)——> фикоцианин (630 нм)——> аллофикоцианин (650 и 670 нм)——> хлорофилл а (670 — 680 нм). О возможности такого рода передачи свидетельствует перекрытие полос флуоресценции и поглощения пигментов в этих рядах; например, антенный хлорофилл а680 имеет максимум флуоресценции, близкий к 700 нм, т. е. в том участке спектра, который в максимальной степени поглощается П7оо {
Реакционные центры. Белки, содержащие длинноволновые формы хлорофиллов (П680 в ФС II и П700 в ФС I) в количестве одна молекула на 200 — 400 молекул других хлорофиллов и способные к первичному фотохимическому разделению зарядов, называют реакционными центрами.
Первичное разделение зарядов в реакционных центрах происходит между молекулами хлорофилловой природы и связано с транспортом электронов. Функцию первичного донора электронов выполняет молекула хлорофилла, находящаяся в синглетном возбужденном состоянии. В реакционном центре ФС II, перешнуровывающем мембрану, первичным донором электронов служит П680, а первичным акцептором — феофетин. В реакционном центре ФС I первичный донор электронов — П700,а акцептор — мономерная форма хлорофилла а695 (А)
Последовательный ход событий в реакционном центре ФС II можно представить в виде следующей схемы:

Порядок реакций в этих процессах определяется продолжительностью того или иного состояния молекул. Молекулы находятся в синглетном возбужденном состоянии 10-10 — 10-9 с. Первичные акцепторы (Фф или A1) получают электроны от своих доноров (П680 или П700 соответственно) за время около 10-12 с. Время обратных рекомбинаций от Фф- на 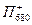 и от
и от 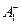 на
на 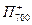 во много раз больше (≈10-6 с). То же самое относится к последующим парам взаимодействующих молекул. Поэтому весь ход реакций резко сдвинут вправо. Z, Пц и QA> A2 — вторичные доноры и акцепторы. Их соответственно окисленное и восстановленное состояние более стабильно, чем состояние окисленных и восстановленных компонентов в реакционных центрах. Таким образом, в реакционных центрах энергия возбужденного
во много раз больше (≈10-6 с). То же самое относится к последующим парам взаимодействующих молекул. Поэтому весь ход реакций резко сдвинут вправо. Z, Пц и QA> A2 — вторичные доноры и акцепторы. Их соответственно окисленное и восстановленное состояние более стабильно, чем состояние окисленных и восстановленных компонентов в реакционных центрах. Таким образом, в реакционных центрах энергия возбужденного
состояния хлорофилла преобразуется в энергию разделенных зарядов, т. е. превращается в химическую энергию (см. рис. 3.4).
Нециклический и циклический транспорт электронов. Для
восстановления одной молекулы NADP+ в процессе фотосинтеза необходимы два электрона и два протона, причем донором электронов является вода. Фотоиндуцированное окисление воды осуществляет ФС II, восстановление NADP+ — фотосистема I. Следовательно, эти две фотосистемы должны функционировать во взаимодействии.
Исходя из данных о квантовом выходе фотосинтеза (необходимы 8 квантов света для выделения одной молекулы О2) из состава компонентов, входящих в ФС I и ФС II, и величин их окислительно-восстановительного потенциала была разработана схема последовательности реакций в световой фазе фотосинтеза (рис. 3.10). Из-за сходства с буквой Z эта схема получила название Z-схемы. Впервые принцип Z-схемы был предложен Р. Хиллом и Ф. Бендаллом (1960) и экспериментально подтвержден работами Л. Дюйзенса (1961). В настоящее время Z-схема, или схема нециклического транспорта электронов при фотосинтезе является общепризнанной и непрерывно пополняется новыми деталями.
В ФС II димер П680, поглотив энергию, эквивалентную 2 квантам коротковолнового красного света, и перейдя в синглетное возбужденное состояние, отдает 2 электрона феофетину (Фф). От Фф электроны, теряя энергию, последовательно передаются на пластохиноны ФС II — QA и QB, на пул липидораство-римых молекул пластохинона (PQ), переносящих через липид-ную фазу мембраны электроны и протоны, на железосерный белок FeSR и цитохром / цитохромного комплекса b6 — ƒ,
восстанавливая Cu-содержащий белок пластоцианин (Пц) (рис. 3.10, табл. 3.1).
Вакантные места («дырки») в П+680 заполняются двумя электронами из содержащего Мп переносчика электронов Z, который в свою очередь восстанавливается с участием системы S. Окисленный белковый комплекс S связывает воду и восстанавливается за счет электронов воды. Для осуществления этой реакции в белковом комплексе S необходимы Мn и С1-, а также Са2+.
Однако описанная цепь реакций резко замедляется, если не возбуждена ФС I, так как в этом случае весь Пц переходит в восстановленное состояние. При возбуждении П700 в реакционном центре ФС I энергией, эквивалентной 2 квантам длинноволнового красного света, 2 электрона захватываются мономерной формой хлорофилла a (A1) и затем последовательно передаются переносчикам электронов А2 и Ав (железосерные белки FeS), ферредоксину (водорастворимый FeS-белок) и ферредоксин: NADP-оксидоредуктазе с FAD в качестве кофактора. Наконец, редуктаза восстанавливает NADP +.

