 2020-01-14
2020-01-14 348
348
В начальную часть кишечника впадают протоки двух пищеварительных желез: печени и поджелудочной железы.
В печени происходит обезвреживание ядовитых веществ, поступающих из кишечника, вырабатывается желчь, эмульгирующая жиры и усиливающая перистальтику кишечника, осуществляется синтез белков и углеводов, накапливаются гликоген, жир, витамины (особенно у акул и тресковых).
У хрящевых рыб печень трёхдольчатая. Масса её достигает 10-20 % массы тела. У многих карповых печень также состоит из трех долей, но у сазана (карпа) их две, а у щуки и окуня одна.
Двухдольчатую печень имеют многие костистые рыбы.
У многих рыб, в частности у карповых, печень включает ткань поджелудочной железы и называется гепатопанкреасом.
Большую роль в пищеварении играют печень и её секрет – желчь. Желчь вырабатывается клетками печени и по дренажной системе желчных протоков поступает в желчный пузырь. В пузыре желчь сгущается до содержания сухого вещества приблизительно 13 %, более половины которого составляют желчные кислоты. Зеленоватый или коричневатый цвет желчи обусловлен присутствием желчных пигментов, представляющих собой продукты деградации гематина. Желчь содержит значительное количество минеральных катионов и анионов – кальция, магния, сульфата, карбоната. В ней содержатся слизистые мукоиды и полярные липоиды, играющие роль смазки для пищевых частиц. В желчи обнаруживается амилазная, липазная и протеазная активность.
Среди веществ желчи основную роль играют поверхностно-активные вещества – желчные кислоты. Они же обусловливают хорошо всем известную горечь желчи. Понижать поверхностное натяжение на границе водной среды и жировых капель способны также желчные пигменты. Поверхностно-активные вещества способствуют распаду жировых капель и стабилизации жировой эмульсии, что увеличивает поверхность пищевого жира, доступную липолитическим ферментам. Кроме того, сверхтонкая эмульсия жира может усваиваться без окончательного гидролиза. Желчные кислоты могут соединяться с высшими жирными кислотами, делая их растворимыми в воде, что улучшает их усвоение. Желчные кислоты, всосавшиеся вместе с жиром, снова попадают в печень и используются повторно.
Желчный пузырь представляет собой как бы боковой карман главного желчного протока. Когда желчный проток перекрыт путем сужения просвета ниже желчного пузыря, желчь накапливается в пузыре, гладкая мускулатура которого в это время расслаблена и сам он растянут. Такое состояние пузыря наблюдается в отсутствие пищеварения. У зимующей молоди карпа желчный пузырь раздут и достигает размеров ягоды смородины. Специфические гормональные воздействия, связанные с питанием и пищеварением, заставляют расслабляться сфинктер желчного протока и напрягаться стенки пузыря. В результате этого желчь попадает в просвет кишечника. Устье желчного протока в кишечнике рыбы определяется по желто-коричневому пятну на отмытой слизистой.
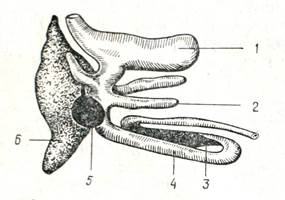
Рис.3 Часть пищеварительного тракта окуня.
1 – желудок; 2 – пилорические придатки; 3 – селезенка; 4 – кишечник; 5 – желчный пузырь; 6 – печень.
Вопрос №3. Что такое раздражимость, возбудимость, возбуждение? Каковы признаки возбуждения? Что такое раздражители? Какие бывают раздражители? Что такое порог раздражения? Что такое торможение?
Нервная система объединяет и согласует деятельность всех систем организма, в результате чего организм способен правильно реагировать на изменение внешней и внутренней среды. Нервная система состоит из центральной и периферической. В центральной нервной системе происходит переработка поступающей информации и принимаются целесообразные решения, а периферическая нервная система передает информацию и точно адресованные команды.
При возбуждении нервного волокна происходит деполяризация, т.е. уменьшение, или инверсия, поляризации мембраны. Нервное возбуждение распространяется по нервам в виде нервных импульсов (спайков), которые характеризуются быстронарастающим и спадающим возбуждением, распространяющимся по нейрону и сопровождающимся изменением физико-химических свойств и электрического потенциала возбужденного участка. Этот процесс является самораспространяющимся от источника возбуждения и происходит он без затухания. Повреждение нервного волокна нарушает проведение импульса. Возбуждение, передающееся по какому-либо нервному волокну, не передается на соседние нервные волокна этого же нервного ствола. Возбуждение по нервному стволу может одновременно передаваться в двух направлениях – центростремительном и центробежном.
Раздражение вагуса ведет к уменьшению сердечного тонуса. Частота сердцебиений не уменьшается, но уменьшается их сила. (Рис.4).

Рис.4. Действие раздражения вагуса на кишечник (а, б, в) и сердце (г).
У беспозвоночных возбуждение в неисчерченных соматических мышцах (как с длинными, так и с короткими волокнами) проводится, вероятно, всегда по нервным волокнам. Широко распространена множественная моторная иннервация мышц, как исчерченных, так и неисчерченных. У позвоночных некоторые неисчерченные мышцы (например, мигательная перепонка) требуют нервного проведения; в других (гладкие мышцы внутренних органов) возбуждение проводится непосредственно от волокна к волокну. О висцеральных мышцах беспозвоночных известно очень мало, но в одной из них (веретенообразная мышца Phascolosoma) возбуждение от волокна к волокну проводится посредством механического растяжения.
Передача возбуждения в нервно-мышечных синапсах осуществляется химическими медиаторами, которые, за исключением ацетилхолина, норадреналина и 5-окситриптомина, еще не идентифицированы. Мембрана нервно-мышечного синапса обычно отличается по своим электрическим свойствам от проводящей мембраны даже той же самой мышечной клетки.
Раздражители, которые воздействуют на механорецепторы, рецепторы равновесия и фонорецепторы, занимают на шкале частот механических колебаний диапазон, простирающийся от постоянного давления через область кратковременных деформаций или низкочастотных вибраций до области ультразвуковых колебаний. Механорецепторы подразделяются на тактильные и проприоцептивные.
Говоря о механическом раздражении, не следует ограничиваться рассмотрением специфических рецепторов, поскольку большинство (а может и все) возбудимых клеток чувствительно к деформации поверхности. Существуют два общих типа механорецепторов: во-первых, те, у которых при механической деформации раздражается часть сенсорного нейрона, и, во-вторых, те, у которых при раздражении в эпителиальных клетках возникают электрические токи, в свою очередь возбуждающие нервные волокна, связанные с этими клетками. В обоих типах рецепторов возбуждение сенсорных аксонов имеет, по-видимому, электрическую природу.
Некоторые механорецепторы проявляют «спонтанную» активность. Следовательно, в центральную нервную систему непрерывно поступают сигналы. В таких органах чувств реакция на раздражитель выражается в изменении прежнего уровня активности, т.е. либо в повышении его, либо в понижении. Это изменение хорошо видно на рецепторах органов равновесия у рыб. Другие механорецепторы не проявляют активности в состоянии покоя. К таким рецепторам относится большинство тактильных волосков, а также имеющие капсулы рецепторы, реагирующие на прикосновение. Механорецепторы независимо от наличия или отсутствия у них спонтанной активности часто находятся под влиянием рефлекторной регуляции со стороны центральной нервной системы.
Мембраны механорецепторов сильно отличаются друг от друга по характеру возбуждения: одни из них в ответ на данное раздражение дают однократный разряд, другие – многократный.
Эхолокацию предметов на расстоянии, по-видимому, применяют некоторые водные насекомые, в небольшой степени рыбы (с помощью органов боковой линии), вероятно, китообразные.
Важным механизмом регуляции активности в нервных центрах служит реципрокное возбуждение и торможение. Один и тот же афферентный путь может быть возбуждающим для одной группы мотонейронов и тормозным для мотонейронов антагонистического действия. Тормозные нейроны, очевидно, действуют, высвобождая какой-то химический передатчик, и один и тот же передатчик может быть тормозным для одного нейрона и возбуждающим для другого.
По-видимому, можно считать всеобщим правилом, что торможение заключается в гиперполяризации, т.е. понижении мембранного потенциала или подавлении возбуждающей деполяризации. В сложных путях рефлекторных реакций происходит отставленное торможение с гораздо большим латентным периодом. Другим механизмом отставленного торможения может служить нейронная цепь с таким распределением процессов во времени, что импульсы, приходящие по определенному пути в соответствующее время, создают рефрактерность для импульсов, приходящих по другим путям.








