 2014-02-12
2014-02-12 802
802Сложный процесс распада ОВ в анаэробных условиях завершают метанобразующие бактерии или метаногены. Они являются облигатными анаэробами, чувствительными к кислороду. Окислительно-восстановительный потенциал среды их роста составляет — 330 мВ и ниже. Среди метанобразующих организмов встречаются психрофильные, мезофильные и термофильные виды. Описаны также галофильные метановые бактерии, обитающие в осадках соленых водоемов. Для обеспечения жизнедеятельности чистых культур метаногенов в первую очередь необходим строжайший анаэробиоз и нейтральная или слабощелочная реакция среды.
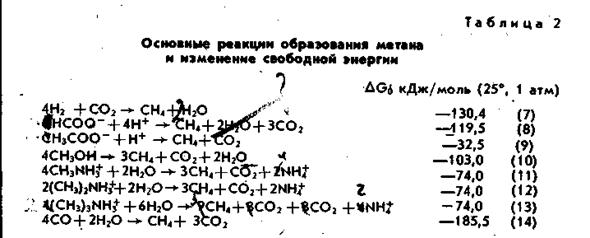 |
Метанобактерии могут использовать в качестве источников энергии и углерода ограниченное количество субстратов— всего 8: СО2+Н2, формиат, закись углерода, метанол, ацетат, моно-, ди- и триметиламины (табл. 2).
Среди них наиболее важным является ацетат, из которого при разложении сложных ОВ образуется более 70% метана (рис. 1). Помимо указанных выше восьми источников углерода никакие другие его соединения не поддерживают рост метанобразующих бактерий, правда имеется сообщение об образование метана из метильных групп многоуглеродных спиртов.
Метаногенные бактерии 90—95% используемого углерода превращают в метан, чтобы за счет сопряженного образования трансмембранного потенциала аккумулировать необходимую для конструктивного обмена энергию, и лишь 5—10% углерода превращается в биомассу. Благодаря указанной особенности до 80—90% ОВ, разлагающегося в процессе развития метаногенного консорциума, превращаются в газ.
По типу использования субстратов все виды метанобразующих организмов могут быть условно разделены на 3 подгруппы (рис. 2). Следует отметить, что здесь мы не рассматриваем группу галофильных метаногенов. Бактерии первой подгруппы используют Н2+СО2, к ним относится большинство метановых бактерий, некоторые из них способны также использовать формиат.
Вторая подгруппа представлена ацетатиспользующими метаногенами. Осуществлять реакцию способны представители только двух родов Methanosarcina и Methanothrix, причем для последних ацетат является единственным субстратом роста, к которому они обладают очень высоким сродством. В настоящее время описано три вида метанотриксов: два мезофильных М. soehngenii и М. concilii и один термофильный М.thermoacetophila.
Так как при реакции выигрыш свободной энергии Гиббса
относительно невелик, то скорость роста бактерий второй
подгруппы достаточно низка — время генерации исчисляется
сутками. Метанотриксы являются определяющей группой
бактерий при очистке сточных вод, они способны использовать ацетат в очень низких концентрациях (до7мкМ) и обеспечивают глубокую очистку стока. |
К третьей подгруппе могут быть отнесены метаносарцины, способные утилизировать все известные к настоящему времени метаногенные субстраты, за исключением формиата. Наиболее предпочтительным субстратом для них является метанол, наименее — ацетат, сродство к которому у них на порядок ниже, чем у метанотриксов. К настоящему времени описано несколько видов метаносарцин, в том, числе термофильная М. thermophila, не использующая водород.
Приведенные в таблице 2 реакции на биохимическом
уровне реализуются с помощью исключительно специфичных только для метаногенов метаболических путей при
участии уникальных метаногенных кофакторов, таких как
метаноптерин, фактор F420, фактор F430, кофермент М и др.
На наличии этих соединений и основан достаточно простой
метод определения водородиспользующих метаногенов в
смешанных культурах с помощью флуоресцентной микроскопии: они дают либо зеленую флуоресценцию за счет
фактора F420 или голубую флуоресценцию за счет 7-метилптерина.
Метанобразующие бактерии представлены видами с раз
ной морфологией. Среди них имеются округлые, ланцето
видные, палочковидные, спиральные, нитевидные и другие
формы. Все метанобразующие бактерии имеют клеточные
стенки. Но в отличие от большинства других бактерий в нее
не входит муреиновая кислота и аминокислоты в D-форме.
У части видов, например, Methanococcus vannieli стенка образована белковыми субъединицами, у других, в том числе
Methanobacterium thermoautotrophicum, содержит особый
пептидглюкан, называемый еще псевдомуреином. В зависимости от состава клеточной стенки метанобразующие бактерии могут окрашиваться как грамположительные или
граммотрицательные бактерии. Особое строение клеточных
стенок обуславливает устойчивость метановых бактерий кантибиотикам, которая варьирует у разных видов. Другое
отличие метанобразующих бактерий от остальных бактерий
проявляется в том, что их клеточные мембраны содержат
глицериновые эфиры полиизопреноидов.








