 2015-03-22
2015-03-22 2729
2729Нижние дыхательные пути в филогенезе формируются как производные элементов жаберного аппарата, а в онтогенезе – как вентральные выросты от пищевода. Однако, в отличие от пищевода (по которому пища проталкивается сокращением стенок), воздуховодные пути должны обеспечивать свободный проход воздуха, то есть зиять, одновременно обеспечивая защиту от попадания пищи. Твёрдую основу гортани составляют хрящевые остатки эмбрионально закладывающихся жаберных дуг, а изменение просвета гортанной щели обеспечивается мышцами, связанными с хрящами и подъязычным аппаратом.
Бесхвостыми амфибиями, некоторыми рептилиями и млекопитающими гортань используется как голосовой аппарат. Для этого производится продавливание воздуха через гортань при суженой гортанной щели. У амфибий звук модулируется при помощи резонирующих мешков, у млекопитающих – голосовых связок и мышц (иногда и резонаторов). Рептилии (кроме гекконов и крокодилов) могут издавать только простые шипящие звуки.
Трахея представляет собой трубку, стенки которой армированы хрящевыми или костными кольцами, полными или нет. Между собой кольца соединены плотной фиброзной соединительной тканью. В результате получается прочная, достаточно гибкая и неспадающаяся трубка. Каудально трахея разделяется на два бронха, имеющих сходное с трахеей строение – это место именуется бифуркацией.
Гортань амфибий разных отрядов имеет разную степень редукции – от набора мелких хрящевых фрагментов до хорошо оформленной гортани из пары черпаловидных хрящей, обрамляющих гортанную щель и перстневидного (кольцевидного) хряща. У бесхвостых амфибий, в связи с короткой шеей и нагнетательным механизмом дыхания, трахеи, фактически, нет. Лёгкие начинаются короткими бронхами непосредственно от гортани. Трахея хвостатых амфибий относительно длинная и может содержать отдельные трахейные хрящи.
Гортанная щель рептилий и птиц обрамлена парными черпаловидными хрящами, ниже располагается, окружая гортань, перстневидный хрящ.
Трахея рептилий образована замкнутыми костными или хрящевыми кольцами, соединёнными плотной соединительной тканью. Вдоль трахеи может лежать полоса губчатого материала – так называемое трахейное лёгкое.
Трахея у птиц, в большинстве случаев, имеет полностью замкнутые кольца. Трахея может превосходить шею и в длину и быть уложена петлей на грудине (и даже в полости киля – у лебедей и журавлей), что придает ей дополнительные резонирующие свойства.
В месте разделения трахеи на два бронха находится нижняя гортань (syrinx) – голосовой аппарат. Она имеет пару внутренних и пару наружных голосовых мембран, натяжение которых регулируется различным количеством голосовых мышц. Голосовые мышцы обеих сторон могут управляться независимо, что позволяет птицам издавать одновременно два разных звука. Звуки издаются птицами и на вдохе и на выдохе. С обеими внутренними мембранами контактирует дивертикул резонирующего межключичного воздушного мешка.
Голосовые мышцы могут крепиться к кольцам трахеи, главных бронхов, или и к тем и к другим. Эти мышцы могут быть редуцированы (страусы), что сильно ограничивает разнообразие издаваемых звуков, а при редукции голосовых мембран (аисты, американские грифы) птицы утрачивают голос вообще.
Хрящевые кольца, составляющие нижнюю гортань, могут окостеневать и частично сливаться в несимметричные образования (у уток).
Гортань млекопитающих состоит из пяти хрящей: пары черпаловидных, перстневидного, щитовидного и надгортанного. Крышу гортани формирует пара черпаловидных хрящей. Их оральные части образованы эластическим хрящом. В момент глотания к ним тесно прилегает свободный край эластического надгортанного хряща (надгортанника). Надгортанник имеет форму изогнутой пластинки, прикреплённой вентрально к гортани и свободной дорсально. Он является новообразованием, производным от складки слизистой оболочки, которая прикрывает гортанную щель у некоторых амфибий и рептилий. Движения надгортанника обеспечиваются сближением при глотании гортани и корня языка с помощью подъязычной мускулатуры. Щитовидный хрящ подстилает гортань вентрально. Внутри гортани имеются голосовые складки, содержащие голосовые связки и мышцы. Между складками – голосовая щель.
Трахея млекопитающих длинная, сформирована хрящевыми кольцами, незамкнутыми дорсально, где проходит трахейная мышца. До разделения на главные бронхи (бифуркации) на правую сторону может отходить непарный трахейный бронх.
ЛЁГКИЕ
Лёгкие - основной (или единственный) орган газообмена у наземных позвоночных. В онтогенезе они (вместе с трахеей) формируются как вентральный, раздваивающийся каудально вырост кишечной трубки. Первоначально лёгкие имеют мешковидную форму и простое строение. Затем приобретают черты и особенности строения, свойственные классу животных.
Лёгкие амфибий часто не играют ведущей роли в процессе газообмена, у представителей некоторых родов они вовсе отсутствуют (безлёгочные саламандры). В этом случае с необходимым объёмом газообмена справляются тонкие, влажные, обильно кровоснабжённые покровы (это особенно актуально для хвостатых амфибий, у которых форма тела обеспечивает б о льшее отношение площади покровов к массе тела), а также слизистая оболочка ротовой полости. У безногих амфибий обычно полное развитие получает только одно лёгкое (как у змей и безногих ящериц). Бесхвостые амфибии имеют два лёгких, которые представляют собой полые мешки. На внутренней поверхности стенок лёгких имеются слабоветвящиеся складки и выросты, увеличивающие её площадь. (Лёгкие амфибий не достигают уровня сложности организации, свойственного даже некоторым двоякодышащим рыбам.)
Механизм лёгочного дыхания амфибий является гулярным, то есть в нём основная роль принадлежит дну ротовой полости и подъязычному аппарату – нагнетающий насос. Механизм дыхания амфибий включает несколько стадий:
-ротовая полость расширяется, её дно опускается, воздух поступает в неё через ноздри и хоаны;
- ноздри закрываются, дно ротовой полости поднимается, воздух проталкивается в лёгкие;
- ротовая полость растягивается при закрытых ноздрях – воздух выходит в неё из лёгких;
- закрывается гортанная щель, воздух выталкивается из ротовой полости наружу через хоаны и ноздри.
У рептилий проявляется тенденция к разделению легких на две функциональные части: в одной собственно происходит газообмен, а другая обеспечивает прокачивание воздуха через первую (максимального выражения это достигает у птиц). Проще всего легкие устроены у большинства ящериц и гаттерий – они сравнительно однородны, и отличаются от легких земноводных лишь сильнее развитыми перегородками и ячейками, обеспечивающими б о льшую функциональную поверхность (по причине отсутствия кожного дыхания).
У крокодилов и черепах дифференцировка на участки для газообмена и для обеспечения вентиляции выражена лучше. У хамелеонов, в отличие от других ящериц, имеются воздушные мешки, выходящие за пределы легких, но газообмен в воздушных мешках не происходит. У примитивных змей (слепозмейки, удавы) левое легкое уменьшено, а у продвинутых змей (ужовые, аспидовые, гадюковые, ямкоголовые) и вовсе отсутствует. У змей настоящее легкое служит именно для прокачивания воздуха, а газообмен происходит в основном в трахейном легком – скоплении губчатой ткани в стенке длинной трахеи.
Механизм дыхания почти всех амниот состоит, как минимум, из изменения положения ребер, что обеспечивает изменение объема полости тела и, следовательно, легких (у млекопитающих добавляется еще и влияние диафрагмы, но у рептилий ее нет). Исключением являются черепахи, поскольку их грудная клетка неподвижна. Наполнение легких воздухом у черепах происходит путём его «заглатывания» при поднимании подъязычного аппарата (как у земноводных), и изменения объема полости тела при поднимании передних конечностей, а также за счёт изменении кривизны мягкой стенки тела возле задних конечностей. Последнее происходит путём сокращения внутренней косой мышцы живота, ставшей инспиратором вместо экспиратора. Последние два механизма настолько важны, что, находясь в защитной позе, черепахи дышать не могут (в определённой мере это компенсируется особенностями сердечно-сосудистой системы черепах).
У двух семейств рептилий, сильно адаптированных к водному образу жизни – трехкоготных черепах и морских змей – в глотке имеются обильно васкуляризированные ворсинки, позволяющие усваивать кислород из воды.
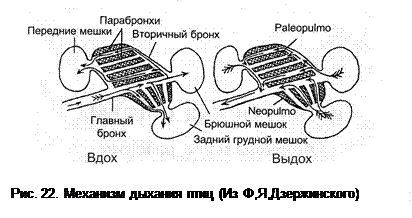
У птиц легкие сквозные, их функциональные элементы – парабронхи – представляют собой трубочки, сквозь которые воздух прокачивается и на вдохе, и на выдохе. Стенки парабронхов продырявлены мелкими отверстиями, сквозь которые воздух проходит в губчатую ткань, окружающую каждый парабронх. Губчатая ткань парабронхов не изолирована и сообщается с соседними парабронхами. В перекладинах губчатой ткани лежат тонкостенные кровеносные капилляры, таким образом, именно здесь происходит газообмен. В отличие от лёгких остальных позвоночных, лёгкие птиц не имеют слепо замкнутых образований – мешков или альвеол, что значительно повышает эффективность дыхания.
Парабронхи связаны с бронхами (более крупными трубками). Бронхи, расположенные исключительно в легких, называются эндобронхами, а выходящие за пределы легких (сообщающиеся с воздушными мешками) – эктобронхами. Легкие у птиц тесно прижаты к дорсолатеральным стенкам грудной полости, так, что рёбра оставляют на них глубокие вдавления. Лёгкие не разделены на доли и лишь снизу контактируют с серозными оболочками. Их объем при вдохе и выдохе не меняется. Морфофункционально легкие птиц делятся на две части – paleopulmo и neopulmo (см. ниже).
Воздушные мешки – гладкостенные, слепозамкнутые образования. Газообмен в них не происходит, но они необходимы для прокачивания воздуха через легкие. С наличием воздушных мешков связано сложное разделение вторичной полости. Объем воздушных мешков непостоянен и связан с изменением объема полости тела. Как у большинства амниот, это совершается за счет изменения положения ребер и грудины. При машущем полете движения крыльев и дыхание синхронизированы, так как часть волокон грудной мышцы крепится на ребрах. У птиц имеются шейные мешки (правый и левый соединены), межключичные (правый и левый соединены), передние грудные, задние грудные и брюшные. Механизм вентиляции легких сводится к следующему (см. рисунок).
При вдохе воздух устремляется во все воздушные мешки, но в брюшные он идет по магистральному бронху, минуя парабронхи лёгкого, а в остальные мешки он идет сквозь лёгкое, по парабронхам, где осуществляется газообмен. (Таким образом, при окончании вдоха в брюшных мешках находится «свежий» воздух, с высоким содержанием кислорода, а в остальных – «отработанный».
При выдохе воздух покидает все воздушные мешки. Из шейных, межключичных и передних грудных мешков на выдохе воздух выходит в главные бронхи и далее в трахею, минуя парабронхи. Из брюшных мешков воздух проходит по парабронхам palaeopulmo, отдавая кислород. Важно, что при этом воздух движется в том же направлении, в котором он проходил из магистрального бронха в шейные, межключичные и передние грудные мешки на вдохе. Однонаправленность потока воздуха на вдохе и выдохе в palaeopulmo обеспечивает высокую эффективность газообмена – птицы могут достигать высоты 9000 метров, недоступной млекопитающим без специального оборудования. Из задних грудных мешков воздух выходит через парабронхи той части легких, что называется neopulmo, в направлении, обратном тому, что было на вдохе. Таким образом, бедный кислородом воздух при обратном движении через парабронхи neopulmo изымает часть кислорода из крови, что снижает эффективность газообмена. Neopulmo усиленно вентилируется у певчих птиц во время пения, что позволяет избежать дыхательного алкалоза при усиленной вентиляции, не соответствующей невысокому потреблению кислорода.
Все воздушные мешки, за исключением грудных, имеют дивертикулы, входящие в скелет или образующие подкожные воздухоносные полости.
Функция воздушных мешков не ограничивается вентиляцией легких. Воздухоносные полости в костях уменьшают плотность тела птиц. Это важно для видов, много времени проводящих в полете. Шейные мешки пневматизируют шейные и грудные позвонки, межключичные – кости плечевого пояса и свободных передних конечностей, брюшные – сложный крестец и кости задних конечностей. Напротив, у ныряющих птиц скелет малопневматичен.
Подкожные воздухоносные полости образуют «подушки безопасности» у птиц, ныряющих в воду с пикирования на большой скорости и способствуют изменению плотности тела для регуляции плавучести у ныряющих птиц. Они надуваются, меняя очертания при токовании и выполняют резонаторную функцию, так как межключичные мешки соприкасаются с внутренними голосовыми мембранами, а также и с другими воздушными мешками. Воздушные мешки осуществляют охлаждение тела птиц (птицы лишены потовых желез). Больше всего тепла вырабатывается в полете при интенсивной работе грудных мышц, а это сопряжено и с усиленной вентиляцией.
Лёгкие млекопитающих имеют гроздевидную внутреннюю структуру. Главные бронхи ветвятся несколько раз, формируя бронхи всё меньшего диаметра – бронхиальное (воздухоносное) дерево. Наконец, от терминальных бронхиол начинается альвеолярное (респираторное) дерево лёгких. В стенках частей респираторного дерева располагаются альвеолы – мешочки с диаметром около 25 мкм, широким входом и очень тонкой стенкой. Стенка альвеолы снаружи окружена эластическими волокнами, к ней тесно прилегают кровеносные капилляры.
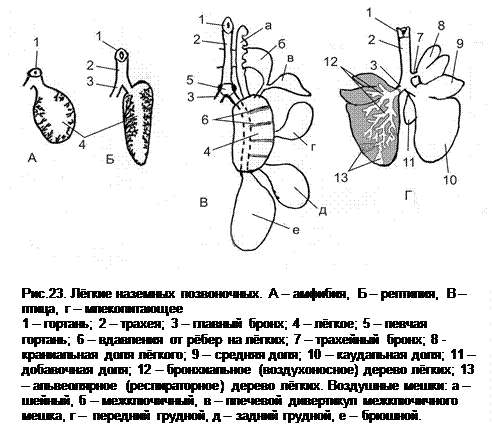
Каждое лёгкое находится в собственной серозной плевральной полости. Париетальный листок плевры выстилает стенки грудной полости, висцеральный – покрывает лёгкое. В плевральной полости содержится минимальное количество плевральной жидкости, отрицательное давление в полости обеспечивает растяжение лёгких вслед за стенками грудной клетки. У млекопитающих наиболее полно используется механизм рёберного дыхания: мышцы-вдыхатели обеспечивают увеличение объёма грудной клетки, сдвигая рёбра краниодорсально. Важную роль также играет появившаяся у млекопитающих диафрагма. Выдох обеспечивается в основном собственной эластической тягой лёгких, мышцы-выдыхатели задействованы в основном при форсированном выдохе (например, при производстве звуков). (Неспадение стенок альвеол обеспечивается наличием особого вещества – сурфактанта, который силой поверхностного натяжения поддерживает альвеолы всегда раскрытыми.) Таким образом, при дыхании лёгкие млекопитающих сильно меняются в объёме. Для более полного использования доступного объёма грудной клетки лёгкие часто сильно рассечены на доли (как правило – это верхушечная, сердечная и диафрагмальньная доли и добавочная на правом лёгком).
Такое строение лёгких делает газообмен в них относительно малопроизводительным, так как даже при выдохе в лёгких остаётся большое количество отработанного воздуха. Установлено, что при спокойном дыхании при каждом цикле воздух в лёгких заменяется примерно на 30%, а во время сна – на 20%.








