 2015-05-10
2015-05-10 927
927Сейчас, просматривая статьи Уотсона и Крика 50-летней давности, удивляешься, как много из предположений оказались верными или близкими к истине – ведь у них не было почти никаких экспериментальных данных. Что касается самих авторов, пятидесятилетие открытия структуры ДНК оба ученых встречают, активно работая теперь уже в разных областях биологии. Дж.Уотсон был одним из инициаторов проекта «Геном человека» и продолжает работать в области молекулярной биологии, а Ф.Крик в начале 2003 г. опубликовал статью о природе сознания.
| Дж.Д. Уотсон, Ф.Г.К. Крик, отдел по изучению молекулярной структуры биологических систем Совета по медицинским исследованиям, Кавендишская лаборатория, Кембридж. 25 апреля, 1953 г. Молекулярная структура нуклеиновых кислот Мы хотим предложить модель структуры соли дезоксирибонуклеиновой кислоты (ДНК). Эта структура обладает новыми свойствами, представляющими интерес для биологии. Структура нуклеиновой кислоты уже предложена Полингом и Кори. Они любезно позволили нам ознакомиться с рукописью их статьи до публикации. Их модель состоит из трех переплетенных цепей с фосфатами, расположенными вблизи оси спирали, и азотистыми основаниями на периферии. По нашему мнению, такая структура неудовлетворительна по двум причинам. Во-первых, мы считаем, что исследуемый материал, дающий рефлексы рентгеновских лучей, является солью, а не свободной кислотой. Без кислотных атомов водорода неясно, какие силы могут поддерживать целостность такой структуры, особенно с учетом того, что отрицательно заряженные фосфатные группы вблизи ее оси будут взаимно отталкиваться. Во-вторых, некоторые из ван-дер-ваальсовых расстояний оказываются слишком малыми. Еще одна трехцепочечная структура предложена Фрейзером (в печати). В его модели фосфаты находятся снаружи, а азотистые основания, соединенные между собой водородными связями, – внутри спирали. В статье эта структура определена очень плохо и по этой причине мы не станем ее комментировать. Мы хотим предложить радикально отличающуюся от этих структуру соли дезоксирибонуклеиновой кислоты. Эта структура состоит из двух спиральных цепей, завитых вокруг общей оси. Мы исходили из обычных предположений, а именно, что каждая цепь образована остатками b-D-дезоксирибофуранозными остатками, соединенными 3',5'-связями. Эти цепи (но не их основания) соединены связями (диадами), перпендикулярными к оси спирали. Обе цепи образуют правую спираль, но, благодаря диадам, имеют противоположные направления. Каждая цепь слегка напоминает модель № 1 Ферберга тем, что основания расположены внутри спирали, а фосфаты снаружи. Конфигурация сахара и атомов вблизи него близка к «стандартной конфигурации» Ферберга, в которой сахар расположен приблизительно перпендикулярно к связанному с ним основанию. Остатки на каждой цепи расположены с шагом 3,4 А по направлению z. Мы предположили, что угол между соседними остатками составляет 36 о, так что эта структура повторяется через каждые 10 остатков, т.е. через 34 А. Расстояние от оси до атома фосфора составляет 10 А. Поскольку фосфаты расположены снаружи, они легко доступны для катионов. Вся структура открыта и содержит довольно много воды. При уменьшении содержания воды можно ожидать, что основания несколько наклонятся, и вся структура станет более компактной. Новым свойством структуры является способ, которым цепи удерживаются друг возле друга за счет пуриновых и пиримидиновых оснований. Плоскости оснований перпендикулярны оси спирали. Они попарно соединены между собой, причем одно основание на первой цепи соединено водородной связью с одним основанием на второй цепи таким образом, что эти основания расположены бок о бок друг с другом и имеют одну и ту же z -координату. Для того, чтобы образовалась связь, одно основание должно быть пуриновым, а другое пиримидиновым. Водородные связи образуются между позицией 1 пурина и позицией 1 пиримидина и между позицией 6 пурина и позицией 6 пиримидина. Предполагается, что основания входят в эту структуру только в наиболее вероятной таутомерной форме (т.е. в кето-, а не в энольной форме). Обнаружено, что только специфические пары оснований могут образовывать связи друг с другом. Эти пары таковы: аденин (пурин) – тимин (пиримидин) и гуанин (пурин) – цитозин (пиримидин). Другими словами, если аденин является одним из членов пары на любой цепи, то в соответствии с этим предположением другим членом пары должен быть тимин. То же относится к гуанину и цитозину. Последовательность оснований на одной цепи, по-видимому, ничем не ограничена. Однако, поскольку могут образовываться только определенные пары оснований, то при заданной последовательности оснований одной цепи последовательность оснований другой цепи определяется автоматически. Экспериментально обнаружено, что в ДНК отношения количества аденинов к количеству тиминов и количества гуанинов к количеству цитозинов всегда близко к единице. Вероятно, невозможно построить такую структуру с рибозой вместо дезоксирибозы, т.к. дополнительный атом кислорода делает ван-дер-ваальсово расстояние слишком малым. Опубликованные до настоящего времени рентгеноструктурные данные по дезоксирибонуклеиновой кислоте недостаточны для строгой проверки нашей модели. Насколько мы можем судить, она приблизительно соответствует экспериментальным данным, но ее нельзя считать доказанной, пока не будет проведено ее сопоставление с более точными экспериментальными данными. Некоторые из них приведены в следующей статье. Нам не были известны детали представленных в ней результатов, когда мы придумывали нашу структуру, которая основывается главным образом, хотя и не только, на опубликованных экспериментальных данных и стереохимических соображениях. Следует заметить, что из постулированного нами специфического образования пар сразу же следует возможный механизм копирования генетического материала. Все детали структуры, включая условия, необходимые для ее построения, и наборы координат атомов будут приведены в последующих публикациях. Мы очень признательны д-ру Джерри Донахью за постоянные советы и критику, особенно относительно межатомных расстояний. Нас также стимулировало общее представление о неопубликованных экспериментальных данных и идеях д-ра М.Г.Ф. Вилкинса и д-ра Р.Э. Франклин и их сотрудников в Кингс-колледже в Лондоне. Один из нас (Дж.Д.У.) получал стипендию Национального фонда детского паралича. |
Еще раз об открытии структуры ДНК
Н.С. Андреева,
доктор физико-математических наук, Институт молекулярной биологии им. В.А. Энгельгардта РАН
В нашей отечественной литературе роль, которую сыграла Розалинда Франклин в становлении молекулярной биологии, в должной степени не отражена. Как правило, открытие структуры ДНК ассоциируется лишь с именами Дж.Уотсона и Ф.Крика, хотя на самом деле три имени должны фигурировать в подзаголовке этой замечательной работы - Уотсон-Крик-Франклин, так как каждый из них внес равноценный, индивидуальный и незаменимый вклад в определение строения вещества наследственности. Но история распорядилась так, что имени Франклин суждено было уйти в небытие *.
* Об этом факте упоминалось недавно в краткой заметке. Подробнее см.: Корочкин Л.И., Фридман С.А. Пионер молекулярной биологии // Природа. 2004. №8. С.75-77.
Все началось с того, что Уотсон и Крик построили свою модель, опираясь на неопубликованные данные Франклин, и в своей знаменитой статье о структуре ДНК не упомянули ее имя [1]. Серьезное и обстоятельное исследование реального вклада Франклин в установление структуры двойной спирали ДНК проведено А.Клугом [2], который опубликовал в 2004 г. большую подборку различных материалов, а также дневники и рабочие тетради Франклин, позволяющие не только понять ее роль в этих исследованиях, но и проследить шаг за шагом, как развивалась мысль и созревало величайшее открытие ХХ в. Мне кажется, что подробное описание этого процесса не только интересно, но и весьма поучительно.
Начнем с исходной ситуации. По сути изучением структуры ДНК занимались три группы. Одна из них, биофизическая, была организована при поддержке Совета по медицинским исследованиям (MRC) сэром Дж.Рэндалом в Лондонском Королевском колледже в 1946 г. “для междисциплинарной атаки на секрет хромосом и близких к ним структур”. Возглавил ее эрудированный и обстоятельный биофизик М.Уилкинс. В этой группе проходил практику студент Р.Гослинг, рентгенографически исследовавший сперму различных животных. Уилкинс преуспел в получении нитей ДНК вытягиванием их из вязких растворов. Нити давали высокое двойное лучепреломление, что свидетельствовало о параллельной ориентации длинных молекул вдоль оси растяжения. Вместе с Гослингом он снял рентгенограмму влажной нити ДНК, которая оказалась чрезвычайно богатой рентгеновскими отражениями (как впоследствии выяснилось, это была рентгенограмма А -формы ДНК).
Именно она вдохновила молодого американского биолога Дж.Уотсона заняться определением структуры ДНК методом рентгеноструктурного анализа. Будь Уотсон более опытным исследователем, он никогда бы не принял подобного решения. Но энтузиазм молодости плюс полное отсутствие знаний в вопросах атомно-молекулярной структуры биополимеров и рентгеноструктурного анализа послужили источником его легкомысленной самоуверенности. Он добился разрешения работать над этой проблемой в знаменитой физической Кавендишской лаборатории, руководимой сэром Лоуренсом Брэггом, основателем рентгеноструктурного анализа. Его приняли в группу М.Перутца, изучавшего структуру гемоглобина и миоглобина.
Как могли Брэгг и Перутц доверить рентгенографическое определение структуры ДНК биологу, не имеющему представления о рентгеноструктурном анализе? Чтобы понять это, необходимо воссоздать атмосферу, царившую в начале 50-х годов в Англии и США в рентгеноструктурных лабораториях, изучающих строение макромолекул. На этот счет имеется много публикаций, в том числе книги Перутца [3, 4], а мне доводилось работать в лаборатории Перутца в конце 50-х и начале 60-х годов и почувствовать эту атмосферу непосредственно.
Помимо основных исследований гемоглобина и миоглобина, Л.Брэгг, М.Перутц и Дж.Кендрью пытались выяснить структуру остова полипептидной цепи, наблюдаемую во множестве фибриллярных белков и, следовательно, отражающую важные принципы строения белковых структур. Эту задачу они решали методом моделирования, в то время очень модным. Именно так были созданы первые модели структуры фибриллярных белков, синтетических полипептидов и, наконец, ДНК, которые впоследствии подтвердились прямыми экспериментальными исследованиями.
Однако когда появился Уотсон, Кембриджская лаборатория потерпела поражение в моделировании структуры фибриллярных -белков. По словам Брэгга, это было самое крупное фиаско в его жизни: модель противоречила законам стереохимии соединений пептидной природы, которые незадолго до этого установил Полинг. В 1951 г. он опубликовал убедительные работы по моделированию структуры остова полипептидной цепи белков, оставив далеко позади модельные построения кембриджских ученых. Труды Полинга, основанные на собственных многолетних исследованиях структуры аминокислот и соединений пептидной природы, позволили ему сформулировать знаменитую теорию резонанса. Опираясь на нее, он показал, что пептидные группы белковых цепей, состоящие из шести атомов - С–СО–NH–C, всегда должны быть плоскими, а структура любого из соединений пептидной природы должна отвечать критерию полного насыщения водородными связями.
Полинг решил построить наиболее выгодную с энергетической точки зрения модель остова полипептидной цепи, состоящего из плоских пептидных групп, и затем проверить ее соответствие экспериментальным данным. Выяснилось, что заданным Полингом критериям полностью отвечает лишь одна структура, названная им -спиралью: она имеет нецелочисленную винтовую ось, т.е. на один ее оборот приходится дробное число пептидных групп (18 в пяти оборотах). Классическая кристаллография отвергала такие винтовые оси, поскольку они не обеспечивают плотного заполнения пространства. Но Полинг искал наиболее выгодную энергетически конформацию остова изолированной полипептидной цепочки, полагая, что к волокнам требования классической кристаллографии применять нельзя. Хотя модель Полинга отвечала всем требованиям стереохимии и выглядела весьма убедительной, но какова должна быть ее рентгенограмма, оставалось неясным. В первой экспериментальной проверке -спирали, проведенной в Кембридже, Перутц выловил на рентгенограммах -белков отражение, подтверждающее модель Полинга. Однако нужны были более обстоятельные эксперименты.
И вот тогда первый раз блеснул своим талантом работавший у Перутца 35-летний физик Ф.Крик. Он вместе с кристаллографом У.Кокреном математически разработал теорию дифракции рентгеновских лучей на спиральных молекулах с винтовыми осями любых порядков, включая нецелочисленные оси. Главное следствие его теории - особое расположение рентгеновских отражений на рентгенограммах спиральных структур. Оно позволяло по виду рентгенограмм идентифицировать спиральную конфигурацию у полимерных молекул в фибриллярных белках и синтетических полипептидах. Расчеты показали, что -фибриллярные белки состоят из скрученных жгутов -спиралей. Стало понятно, что законы погасаний рентгеновских отражений от структур с винтовыми осями, которые использует классическая кристаллография, - лишь частный случай общей теории Крика [5].
Конечно, эта работа придала Крику солидный запас уверенности в себе, и когда появился Уотсон с навязчивой идеей определить структуру ДНК, Крик, видимо, решил и здесь попробовать свои силы. Уязвленный неудачей в построении модели -фибриллярных белков, Брэгг дал молодым ученым свое согласие. Вдохновленные успехом Полинга и кажущейся легкостью в построении модели полипептидной цепи белков, Уотсон и Крик намеревались таким же способом определить строение ДНК. Рентгеноструктурные данные, полученные еще в 40-е годы В.Астбери, говорили, что ДНК свойственна определенная стабильная и упорядоченная структура. Никакой другой информацией Уотсон и Крик на первых порах не располагали.
Если при моделировании -спирали речь шла о плотной упаковке плоских пептидных групп, состоявших из шести атомов, с точно установленным расположением, то отнюдь не плоские нуклеотидные группы содержали значительно больше атомов. Об их расположении можно было лишь гадать, да кроме того, было вообще неизвестно, из скольких цепей состоит молекула. Поэтому для определения структуры ДНК требовалась иная схема, противоположная той, которая реализовалась при анализе структуры фибриллярных белков (когда сперва строилась модель, а потом она проверялась экспериментально). Здесь в первую очередь оказались нужны независимые экспериментальные данные о параметрах структуры. Уотсон и Крик были абсолютными новичками в изучении стереохимии нуклеотидных соединений. Так что, в отличие от Полинга, никаких основополагающих идей о том, какой должна быть структура ДНК, у них не было, кроме предположения Крика, что это спираль. Отсюда становится ясным значение результатов экспериментальных исследований как исходного пункта для построения правильной модели ДНК.
В 1950 г. Рэндал решил усилить рентгеноструктурные исследования ДНК в Королевском колледже, пригласив физико-химика Розалинду Франклин. Ее рентгенографические исследования аморфных углей, требующие экспериментального мастерства и серьезной математической подготовки в области теории рассеяния рентгеновских лучей, снискали ей высокую репутацию среди коллег. Поскольку Уилкинс как руководитель был совершенно не нужен Франклин, оба стали заниматься изучением структуры ДНК отдельно.
Существенно улучшив способ получения ориентированных препаратов ДНК и условия съемки рентгенограмм, Розалинда сразу обнаружила две структурные формы у ДНК - кристаллическую А -форму и паракристаллическую В -форму - и показала, что свои данные Астбери получил от смеси этих двух форм. Свое внимание она сосредоточила на кристаллической А -форме ДНК, пытаясь по плотности вещества и объему ячейки определить количество цепей, проходящих через одну элементарную ячейку.
Тем временем Уотсон и Крик почти вслепую начали собирать модель ДНК из латунных стержней, изготавливаемых в мастерских Кавендишской лаборатории *. Побывав в Королевском колледже на кристаллографическом семинаре Франклин, биолог Уотсон не вынес ничего, кроме того, что молекула ДНК может состоять из трех цепей. На самом деле Розалинда подчеркивала, что если учесть молекулы воды в элементарной ячейке кристаллической формы, то масса вещества в ячейке соответствует скорее двум цепям ДНК. Ничтоже сумняшеся, Уотсон и Крик принялись строить модель спиральной структуры ДНК из трех цепей с переплетающимися фосфатно-сахарными остовами вблизи оси цилиндрической молекулы и торчащими наружу основаниями, по примеру того, что наблюдается в -спиральных белках, где нерегулярно чередующиеся боковые группы торчат наружу. Как же иначе можно было расположить нерегулярно чередующиеся азотистые основания? Построив в течение одной недели модель, Уотсон и Крик пригласили Франклин. Она сразу спросила их, куда они девали воду, а потом разнесла модель в пух и прах как не отвечающую экспериментальным данным. Услышав об этом, Брэгг категорически запретил Уотсону и Крику заниматься играми в ДНК в его лаборатории, и они прервали свою работу на довольно продолжительное время.
* Стержневые модели атомов представляли собой сделанные в определенном масштабе рогатки из расходящихся из одной точки стерженьков под углами, соответствующими валентным углам атомов, с длинами, пропорциональными длинам валентных связей.
Надо сказать, что Уотсон и Крик были не одиноки в своей ошибке. Такую же модель построил и Полинг. Но он сделал еще более грубую ошибку, расположив заряженные фосфатные группы совсем близко друг от друга, не объяснив, почему они не отталкиваются. Ну ведь у Полинга не было экспериментальных данных о ДНК, как, впрочем, и у Уотсона с Криком, если не считать то, что постиг Уотсон на семинаре у Франклин.
Тем временем Розалинда продолжала свои эксперименты. Она получила совершенно замечательную рентгенограмму В -формы ДНК, которая сейчас называется канонической и приводится во многих учебниках [6]. Самым важным в этой рентгенограмме было расположение отражений, четко соответствующее спиральной структуре молекулы, с 10 идентичными группами в одном обороте спирали, равном 34 A и с шагом спирали в 3.4 A. Рентгенограммы свидетельствовали также, что диаметр цилиндрической молекулы ДНК равен приблизительно 20 A. Получив эту рентгенограмму, Розалинда замешкалась. Она никак не могла понять, как соотносится структура В -формы ДНК со структурой кристаллической А -формы, на рентгенограммах которой видимых признаков спиральной формы молекулы ДНК не наблюдалось.
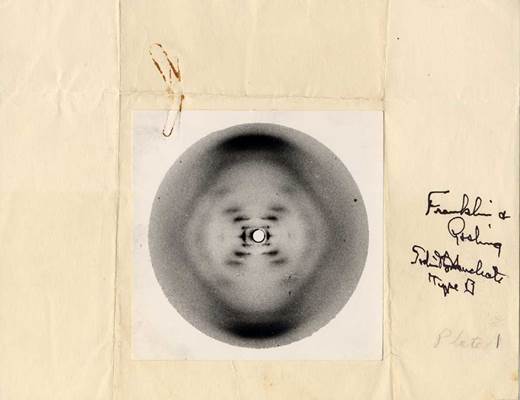
Рентгеновская дифракция на В-форме тимусной ДНК. Снимок Р. Франклин и Р. Гослинга
Согласно теории рассеяния рентгеновских лучей на цепных молекулах, это различие находит вполне четкое объяснение. Не углубляясь в дебри специальных рассуждений, скажу лишь, что в 1950-1953 гг., будучи аспиранткой кафедры рентгеноструктурного анализа физического факультета МГУ, я занималась рентгеноструктурным анализом фибриллярных белков. Работ Розалинды Франклин я не знала, но работу Крика по дифракции на спиралях изучила до дыр. Крик рассчитывал рассеяние рентгеновских лучей изолированной цепной молекулой, не учитывая влияние взаимной упаковки молекул на дифракционную картину. В паракристаллической В- форме ДНК, где молекулы окружены водной шубой, соседние молекулы не могут находиться в регистре друг относительно друга, они всего лишь параллельны, но могут быть повернуты на любой угол вокруг своей длины и смещены вдоль нее относительно друг друга любым образом, т.е. пространственной кристаллической решетки нет. Лишь в плоскости, перпендикулярной длине молекул, они образуют двумерную квазирешетку, так как расстояние между цепями выдерживается не очень строго. Поэтому дифракционная картина от паракристаллических фибриллярных структур определяется в основном независимым рассеянием молекул, для которых полностью применима теория Крика.
Взаимная упаковка молекул проявляется лишь в виде максимумов на экваторе рентгенограммы. В то же время при дифракции рентгеновских лучей от кристаллических полимерных структур со спиральными молекулами упаковка молекул вызывает погасание большей части рассеянного излучения, оставляя лишь точечные рефлексы, обусловленные кристаллической решеткой. При этом картина рассеяния рентгеновских лучей изолированными спиральными молекулами оказывается затушеванной. Видимо, это обстоятельство не пришло на ум Розалинде. Трудности усугублялись тем, что спираль в А -форме немного отличается от спирали в В -форме ДНК, поэтому Франклин не торопилась комбинировать данные, полученные для разных форм.

Фрагмент странички из публикации в "Nature" (Апрель 1953 г.) [6]
Вся страничка (320 Кб)
Уотсон и Крик ознакомились с моделью Полинга по рукописи, привезенной в Кембридж сыном Полинга, поступающим в аспирантуру к Перутцу. Уотсон повез этот текст Уилкинсу, который в ответ показал копию рентгенограммы В -формы ДНК, полученной Франклин. Уотсон, к тому времени поднаторевший в рентгеноструктурном анализе, немедленно передал информацию Крику. Можно себе представить, как обрадовался Крик, узнав о прекрасном воплощении своей теории на рентгенограмме ДНК. Возможность построения правильной модели ДНК вновь забрезжила; Уотсон и Крик бросились к Брэггу с просьбой продолжить свои построения. Брэгг, узнав об ошибке Полинга, такое разрешение дал.
Надо полагать, что энтузиазм Уотсона и Крика в то время нарастал, и в этой атмосфере Перутц ознакомил их с подробным годовым отчетом Розалинды Франклин о проведенных ею исследованиях структуры ДНК. По сути отчет содержал весь комплекс экспериментальных данных, необходимых для построения модели ДНК. Однако Франклин считала, что данные, полученные для одной формы, пока преждевременно переносить на другую, и поэтому медлила с публикацией своих экспериментальных работ. О том, что ее результаты попали в руки Уотсона и Крика, она не знала.

Розалинда Франклин (1920-1958)
Следует отдать должное тому, как быстро и талантливо Крик скомбинировал данные Франклин для построения единственно возможной и правильной модели молекулы ДНК. Он понял, что спираль состоит из двух коаксиальных спиралей, образованных фосфатно-сахарными остовами цепей и направленных, в соответствии с требованиями симметрии С2, найденной в А -форме, в противоположные стороны. Теперь можно было приступать к практическому построению вполне добротной модели остова молекулы ДНК.
Строил модель Уотсон, а Крик им руководил. Можно ли сказать, что модель Уотсона-Крика оказалась простой иллюстрацией рентгеноструктурных данных, полученных Франклин? И да, и нет, потому что это может касаться лишь той части модели, которая представляет строение фосфатно-сахарного каркаса молекулы; кстати, это именно двойная спираль. Но данных Франклин было недостаточно, чтобы правильно расположить азотистые основания внутри двойной спирали, хотя они и указывали, что основания, будучи плоскими и имея толщину около 3.4 A, должны лежать стопкой перпендикулярно ее оси [7].
Работа, связанная с размещением плоских азотистых оснований внутри молекулы, стала результатом творческого поиска Уотсона и Крика, главным образом Уотсона. Посторонние структурные данные, использованные ими, сводились лишь к правильным таутомерным формам оснований, найденным Дж.Доною, что сыграло немаловажную роль в построении правильной модели. Но основным залогом успеха этой части работы была правильная модель фосфатно-сахарного остова, которая неукоснительно вела Уотсона и Крика к их великому открытию.
Эта модель подсказала, что плоские основания двух противоположно направленных цепей, присоединенные к их сахарным группам, которые располагаются на одном уровне по высоте спирали, должны лежать точно в одной плоскости. Следовательно, они должны образовывать пары, но как в них основания располагаются друг относительно друга?
Задача решалась перебором различных пар оснований. Сразу были отброшены два неприемлемых варианта: пары пурин-пурин просто не влезали во внутреннюю полость цилиндра, а пары пиримидин-пиримидин были настолько малы, что проваливались, не цепляясь одновременно за оба крючка гликозидных связей двух противоположно направленных цепей. Судя по всему, немалую роль в быстроте этого поиска сыграла работа биохимика Э.Чаргаффа о равенстве содержания аденина и тимина, а также гуанина и цитозина во всех препаратах ДНК [8]. И действительно, пары аденин-тимин и гуанин-цитозин как по выкройке размещались во внутренней полости цилиндра, и их гликозидные связи точно попадали в модели на нужное место. Вместе с тем соприкасающиеся поверхности оснований в этих парах отличались высокой комплементарностью, и при этом насыщались водородные связи, что не могло быть случайным. Вероятнее всего, открытие Чаргаффа оплодотворило работу Уотсона и Крика по моделированию расположения оснований в спирали ДНК.
Итак, в единственно приемлемой со стереохимической точки зрения структуре ДНК, отвечающей всем экспериментальным данным, последовательность оснований в одной цепи должна обязательно быть комплементарна последовательности оснований в другой. Иначе будут возникать нерегулярные пустоты и искривления цепочек, из-за чего резко упадет стабильность всей структуры. Вместе с тем на последовательность оснований в одной из цепей найденная структура никаких ограничений не налагала. Это также означало, что информация об этой последовательности может надежно храниться внутри молекулы. Следовательно, генетическая информация должна записываться именно в виде последовательности оснований, которая может передаваться по наследству, в случае, если в процессе репликации ДНК спираль расплетается, и одна из цепей служит шаблоном для биосинтеза другой.
Наверное, счастье, которое испытали Уотсон и Крик, поняв замечательные потенции найденной ими структуры, было не меньшим, чем при присуждении им Нобелевской премии. Казалось бы, что на фоне такого удивительного подарка судьбы можно быть более щедрыми и отдать Розалинде Франклин то, что ей принадлежало по праву, т.е. отметить, что ее работы послужили экспериментальной основой модели. Но, увы, этого не произошло: о том, что ее данные позволили им построить правильную модель ДНК, не сказано ни слова, и ее имя вообще не упомянуто в их знаменитой статье, опубликованной в “Nature” в апреле 1953 г. Правда, в своей книге “Двойная спираль”, вышедшей спустя 15 лет, Уотсон вспоминает Розалинду Франклин [9]. Приходится лишь удивляться контрасту между совершенством Природы, выразившимся в удивительно гармоничной структуре молекулы ДНК, и ограниченностью человеческой натуры, проявившейся при ее открытии. Вскоре после этих событий Розалинда Франклин умерла от рака. Ей было всего 37 лет.
Кратко резюмируя изложенное, можно сказать что в учебных пособиях по биологии, равно как и в лекциях на соответствующую тему, правильнее говорить не о двойной спирали ДНК Уотсона и Крика, а о двойной спирали ДНК Уотсона, Крика и Франклин.
| Еврейская мама ДНК |
 Празднуя "пятидесятилетний юбилей" пресловутой двойной спирали, не будем забывать о заслугах молекулярного биолога Розалинд Фрэнклин. В марте 1953 года Морис Уилкинс, сотрудник лондонского Кингз-Колледжа, написал своему другу Фрэнсису Крику, работавшему в "лагере противника" - Кавендишской лаборатории: "на следующей неделе наша смуглая леди нас покидает". Имелось в виду, что коллега Уилкинса, Розалинд Фрэнклин, увольняется. К этому времени Крик и Джеймс Уотсон уже завершили работу над моделью структуры молекулы ДНК, позднее прозванной "двойная спираль". При этом они воспользовались данными исследований Фрэнклин, без ее ведома полученными от Уилкинса. Через месяц Уотсон и Крик опубликовали свою модель в журнале "Нейчер". За это революционное открытие, подсказавшее ключ к тайне копирования генетической информации, Уотсон, Крик и Уилкинс в 1962 году получили Нобелевскую премию в области медицины. За четыре года до их триумфа 37-летняя Фрэнклин умерла от рака яичников, так и не узнав о своем крупном вкладе в одно из величайших научных достижений нашей эпохи. Сейчас, когда на всем земном шаре проходят торжества по случаю 50-летия со дня открытия структуры ДНК, Фрэнклин наконец-то удостоилась кое-каких посмертных почестей. Скорее всего, вклад Фрэнклин так и остался бы непризнанным широкой общественностью, если бы не невольная помощь Уотсона. Его книга мемуаров "Двойная спираль", опубликованная в 1968 году и вошедшая в списки бестселлеров, буквально переполнена нелестными отзывами о Фрэнклин, одной из немногих женщин √ более того, одной из немногих женщин еврейского происхождения, √ которые занимались молекулярной биологией и кристаллографией. Но именно благодаря воспоминаниям Уотсона Фрэнклин вошла в "пантеон святых" феминизма. "Если бы я дала выход своим бунтарским порывам, то обязательно предложила бы переименовать модель ДНК √ пусть это будет модель Уотсона-Крика-Фрэнклин", √ говорит Линн Осмэн Элкин, преподаватель биологических наук из университета штата Калифорния, что в Хейуорде. Линн Элкин, а также ряд знаменитых ученых, когда-то сотрудничавших с Фрэнклин, частвовали в съемках фильма "Тайна фото 51", который будет показан 22 апреля на канале "Пи-би-эс" в рамках научно-популярного тележурнала "NOVA". В роли рассказчика выступает актриса Сигурни Уивер. Режиссер Гэри Глассмэн снял этот документальный фильм по мотивам книги Бренды Мэддокс "Розалинд Фрэнклин: Смуглая леди ДНК", выпущенной в 2002 году издательством "ХарперКоллинз". Название фильма отсылает к уникальному рентгеновскому снимку молекулы ДНК, сделанному Фрэнклин, - он-то и подсказал Уотсону разгадку тайны двойной спирали. Рассказывая о Фрэнклин, авторы фильма уделяют главное внимание ее научной деятельности. Подчеркивается, "какой вопиющей несправедливостью было непризнание ее вклада в описание структуры ДНК", √ сообщил нашему корреспонденту Глассмэн по телефону из Род-Айленда. Фильм вскрывает причины произошедшего: среди них √ и психологическая несовместимость Фрэнклин и Уилкинса, и тайные интриги в научном сообществе, и классовая вражда, и гендерно-религиозные предрассудки. Словом, вина лежит на патриархальном духе Кингз-Колледжа, где все решала клика "своих людей". Интервью с некоторыми из коллег Фрэнклин √ в том числе кристаллографом, нобелевским лауреатом Аароном Клугом, √ свидетельствуют, что Фрэнклин была одним из виднейших специалистов в области рентгеновской кристаллографии, внесла крупный вклад в науку и добилась блестящих успехов. В своей биографии Мэддокс яркими красками обрисовывает характер и деятельность Фрэнклин: перед нами предстает женщина, которая была самоотверженным и дотошным исследователем, обожала Францию (три года проработала по специальности в Париже), занималась альпинизмом. Что до национального самоощущения, то Фрэнклин считала себя еврейкой и гордилась этим. "Я всегда обращаю особое внимание на детство и воспитание, как люди становятся такими, какими мы их теперь знаем", - сказала Мэддокс по телефону из Лондона нашему корреспонденту. На взгляд Мэддокс, еврейские корни Фрэнклин были одним из основных факторов, повлиявших как на становление ее характера, так и на ее путь в науке. 380-страничная биография открывается занимательным рассказом о жизни английских евреев, а также о предках Фрэнклин, переехавших в Великобританию из силезского города Бреслау (там они носили фамилию Френкель). Кстати, эта семья ведет свою родословную от самого царя Давида. Английские евреи, пишет Мэддокс, были "большими англичанами, чем сами англичане. Они переодевались к обеду, стремились быть представленными к королевскому двору┘ Многие праздновали не только Песах, но и Рождество, ели кошерную пищу, что не мешало им играть в крикет". Большим авторитетом в семье пользовался Артур Фрэнклин, приходившийся Розалинд дедом. Фрэнклин находилась под сильным влиянием своего деда. Фрэнклины активно занимались общественной деятельностью и сознавали свой долг перед соплеменниками (так, согласно завещанию Артура Фрэнклина, его наследниками могли стать лишь те, кто состоит в браке с евреями и еврейками). Двоюродный дед Фрэнклин, Герберт Сэмюэль, был видным дипломатом - первым высоким комиссаром Палестины и автором меморандума, который повлек за собой Декларацию Бальфура. Хотя Фрэнклин никогда не сталкивалась с открытыми проявлениями антисемитизма, вполне возможно, что она всегда ощущала некоторый холодок со стороны своих британских коллег-неевреев. "Ее еврейское происхождение, безусловно, было дополнительным препятствием для признания Фрэнклин полноправным соавтором в обществе, где власть принадлежала мужчинам", - говорит Глассмэн. А затем, подхватывая одну из главных тем книги Мэддокс, отмечает, что идея самоотверженного служения науке и человечеству была "привита Фрэнклин с детства" благодаря еврейской среде, в которой она выросла. Возражая отцу, верившему в загробную жизнь, Фрэнклин √ на тот момент студентка Кембриджа √ писала: "Я согласна с тем, что вера √ непременное условие для того, чтобы состояться в жизни. На мой взгляд, все, что нужно для веры, - это убежденность в том, что к цели мы обязательно приблизимся, если только постараемся, а также в том, что нашей цели (улучшения жизни человечества в настоящем и будущем) действительно стоит добиваться". Фильм "Тайна фото 51" будет впервые показан 22 апреля в 8 часов вечера по Восточному времени (расхождение с Москвой 7 часов. √ Пер.) на канале "Пи-би-эс". Джулия Голдмэн Перевод Светланы Силаковой, sem40 21.04.2003 Празднуя "пятидесятилетний юбилей" пресловутой двойной спирали, не будем забывать о заслугах молекулярного биолога Розалинд Фрэнклин. В марте 1953 года Морис Уилкинс, сотрудник лондонского Кингз-Колледжа, написал своему другу Фрэнсису Крику, работавшему в "лагере противника" - Кавендишской лаборатории: "на следующей неделе наша смуглая леди нас покидает". Имелось в виду, что коллега Уилкинса, Розалинд Фрэнклин, увольняется. К этому времени Крик и Джеймс Уотсон уже завершили работу над моделью структуры молекулы ДНК, позднее прозванной "двойная спираль". При этом они воспользовались данными исследований Фрэнклин, без ее ведома полученными от Уилкинса. Через месяц Уотсон и Крик опубликовали свою модель в журнале "Нейчер". За это революционное открытие, подсказавшее ключ к тайне копирования генетической информации, Уотсон, Крик и Уилкинс в 1962 году получили Нобелевскую премию в области медицины. За четыре года до их триумфа 37-летняя Фрэнклин умерла от рака яичников, так и не узнав о своем крупном вкладе в одно из величайших научных достижений нашей эпохи. Сейчас, когда на всем земном шаре проходят торжества по случаю 50-летия со дня открытия структуры ДНК, Фрэнклин наконец-то удостоилась кое-каких посмертных почестей. Скорее всего, вклад Фрэнклин так и остался бы непризнанным широкой общественностью, если бы не невольная помощь Уотсона. Его книга мемуаров "Двойная спираль", опубликованная в 1968 году и вошедшая в списки бестселлеров, буквально переполнена нелестными отзывами о Фрэнклин, одной из немногих женщин √ более того, одной из немногих женщин еврейского происхождения, √ которые занимались молекулярной биологией и кристаллографией. Но именно благодаря воспоминаниям Уотсона Фрэнклин вошла в "пантеон святых" феминизма. "Если бы я дала выход своим бунтарским порывам, то обязательно предложила бы переименовать модель ДНК √ пусть это будет модель Уотсона-Крика-Фрэнклин", √ говорит Линн Осмэн Элкин, преподаватель биологических наук из университета штата Калифорния, что в Хейуорде. Линн Элкин, а также ряд знаменитых ученых, когда-то сотрудничавших с Фрэнклин, частвовали в съемках фильма "Тайна фото 51", который будет показан 22 апреля на канале "Пи-би-эс" в рамках научно-популярного тележурнала "NOVA". В роли рассказчика выступает актриса Сигурни Уивер. Режиссер Гэри Глассмэн снял этот документальный фильм по мотивам книги Бренды Мэддокс "Розалинд Фрэнклин: Смуглая леди ДНК", выпущенной в 2002 году издательством "ХарперКоллинз". Название фильма отсылает к уникальному рентгеновскому снимку молекулы ДНК, сделанному Фрэнклин, - он-то и подсказал Уотсону разгадку тайны двойной спирали. Рассказывая о Фрэнклин, авторы фильма уделяют главное внимание ее научной деятельности. Подчеркивается, "какой вопиющей несправедливостью было непризнание ее вклада в описание структуры ДНК", √ сообщил нашему корреспонденту Глассмэн по телефону из Род-Айленда. Фильм вскрывает причины произошедшего: среди них √ и психологическая несовместимость Фрэнклин и Уилкинса, и тайные интриги в научном сообществе, и классовая вражда, и гендерно-религиозные предрассудки. Словом, вина лежит на патриархальном духе Кингз-Колледжа, где все решала клика "своих людей". Интервью с некоторыми из коллег Фрэнклин √ в том числе кристаллографом, нобелевским лауреатом Аароном Клугом, √ свидетельствуют, что Фрэнклин была одним из виднейших специалистов в области рентгеновской кристаллографии, внесла крупный вклад в науку и добилась блестящих успехов. В своей биографии Мэддокс яркими красками обрисовывает характер и деятельность Фрэнклин: перед нами предстает женщина, которая была самоотверженным и дотошным исследователем, обожала Францию (три года проработала по специальности в Париже), занималась альпинизмом. Что до национального самоощущения, то Фрэнклин считала себя еврейкой и гордилась этим. "Я всегда обращаю особое внимание на детство и воспитание, как люди становятся такими, какими мы их теперь знаем", - сказала Мэддокс по телефону из Лондона нашему корреспонденту. На взгляд Мэддокс, еврейские корни Фрэнклин были одним из основных факторов, повлиявших как на становление ее характера, так и на ее путь в науке. 380-страничная биография открывается занимательным рассказом о жизни английских евреев, а также о предках Фрэнклин, переехавших в Великобританию из силезского города Бреслау (там они носили фамилию Френкель). Кстати, эта семья ведет свою родословную от самого царя Давида. Английские евреи, пишет Мэддокс, были "большими англичанами, чем сами англичане. Они переодевались к обеду, стремились быть представленными к королевскому двору┘ Многие праздновали не только Песах, но и Рождество, ели кошерную пищу, что не мешало им играть в крикет". Большим авторитетом в семье пользовался Артур Фрэнклин, приходившийся Розалинд дедом. Фрэнклин находилась под сильным влиянием своего деда. Фрэнклины активно занимались общественной деятельностью и сознавали свой долг перед соплеменниками (так, согласно завещанию Артура Фрэнклина, его наследниками могли стать лишь те, кто состоит в браке с евреями и еврейками). Двоюродный дед Фрэнклин, Герберт Сэмюэль, был видным дипломатом - первым высоким комиссаром Палестины и автором меморандума, который повлек за собой Декларацию Бальфура. Хотя Фрэнклин никогда не сталкивалась с открытыми проявлениями антисемитизма, вполне возможно, что она всегда ощущала некоторый холодок со стороны своих британских коллег-неевреев. "Ее еврейское происхождение, безусловно, было дополнительным препятствием для признания Фрэнклин полноправным соавтором в обществе, где власть принадлежала мужчинам", - говорит Глассмэн. А затем, подхватывая одну из главных тем книги Мэддокс, отмечает, что идея самоотверженного служения науке и человечеству была "привита Фрэнклин с детства" благодаря еврейской среде, в которой она выросла. Возражая отцу, верившему в загробную жизнь, Фрэнклин √ на тот момент студентка Кембриджа √ писала: "Я согласна с тем, что вера √ непременное условие для того, чтобы состояться в жизни. На мой взгляд, все, что нужно для веры, - это убежденность в том, что к цели мы обязательно приблизимся, если только постараемся, а также в том, что нашей цели (улучшения жизни человечества в настоящем и будущем) действительно стоит добиваться". Фильм "Тайна фото 51" будет впервые показан 22 апреля в 8 часов вечера по Восточному времени (расхождение с Москвой 7 часов. √ Пер.) на канале "Пи-би-эс". Джулия Голдмэн Перевод Светланы Силаковой, sem40 21.04.2003 |
Джеймс Уотсон (James Dewey Watson) родился в городе Чикаго в 1928 году. В 1947 году получил степень бакалавра в Чикагском университете, в 1950 году — степень доктора наук в Индианском университете в Блумингтоне, обе степени по специализации зоология. После проведения исследований в Копенгагене по Национальному исследовательскому гранту он получил грант Национального фонда детского паралича для проведения исследований в Кембридже (Англия), где вместе с Фрэнсисом Криком открыл структуру дезоксирибонуклеиновой кислоты (ДНК). Уотсон и Крик предположили, что молекула ДНК имеет форму двойной винтовой спирали. Уотсон, Крик и Морис Уилкинc, подтвердившие структуру ДНК с помощью рентгенокристаллографии в Королевском колледже (King's College) в Лондоне, в 1962 году получили Нобелевскую премию в области физиологии и медицины за свое открытие.
После работы в Кембридже Уотсон провел два года в Калифорнийском технологическом институте, затем приступил к работе на биологическом факультете Гарвардского университета, а в 1961 году стал профессором. В 1968 году, работая в Гарварде, он становится директором Лаборатории количественной биологии в Колд-Спринг-Харбор (Лонг-Айленд, Нью-Йорк). В 1976 году Уотсон оставил свой пост в Гарварде и стал штатным директором Лаборатории Колд-Спринг-Харбор (Cold Spring Harbor Laboratory). Под его руководством прославленное, но испытывающее финансовые проблемы учреждение получило вторую жизнь. Уотсон направил энергию ученых лаборатории в область вирологии опухолей, из которой исходит сегодняшнее понимание онкогенов (раковых генов) и молекулярной основы рака. В 1994 году он стал президентом Лаборатории.
В 1988 году Уотсон был назначен заместителем директора исследования генома человека, проводимого Национальным институтом здоровья (National Institutes of Health), а в 1989 году — директором Национального центра исследования генома человека при Национальном институте здоровья. В 1992 году Уотсон оставил свой пост в Национальном центре исследования генома человека после успешного проведения всемирных исследований по картированию и секвенированию генома человека.
Джеймс Д. Уотсон является членом Американского Биохимического Общества (1958) и Американской Ассоциации Исследований рака (1972), а также почетным членом Американской Академии Искусств и Наук (1958), Национальной Академии Наук (1962), Датской Академии Искусств и Наук (1963), Колледжа Клэр в Кембриджском Университете (1968), Американского Общества Философии (1977), Клуба Атенеум в Лондоне (1980), Королевского Общества в Лондоне (1981), Российской Академии Наук (1989), Оксфордского Университета (1994), Украинской Национальной Академии Наук (1995), Университетского Колледжа Галуэя, Общества Пуритан и Ученых (1995), Лондонского Института Биологии (1995), Института Фундаментальных Исследований Тата (1996), Королевского Общества Эдинбурга (1999), а также Национальной Академии Наук Индии (2001).
Джеймс Д. Уотсон награжден огромным количеством наград, получил почетные степени 32 университетов и опубликовал 9 книг: «Молекулярная биология гена» (1965, 1970, 1976, 1987), «Двойная спираль» (1968), «История ДНК» (1981), «Молекулярная биология клетки» (1983, 1989, 1994), «Рекомбинантная ДНК: краткий курс» (1983, 1992), «Страсть к ДНК» (2000), «Гены, девушки и Гамов: после двойной спирали» (2002), «ДНК: секрет жизни» (2003), «Избегайте скучных людей: уроки из жизни в науке» (2007).








