 2015-10-16
2015-10-16 5293
5293Организация генома
Исследование эволюции и построение гипотез относительно механизмов эволюционных изменений в настоящее время трудно представить без привлечения генетики. В начале двадцатого века именно использование генетических методов обеспечило прорыв в эволюционных исследованиях и подъём их на качественно новый уровень.
Эволюционные концепции Ламарка и Дарвина базировались на исследовании внешних признаков и свойств живых организмов, предположения о механизме их наследования носили умозрительный характер. Представление о материальных носителях наследственности сводилось к принципу непрерывности, аналогу жидкости, способной к бесконечному разбавлению, несмотря на известные достаточно красноречивые факты, явно указывающие на дискретный характер наследования некоторых признаков по хорошо изученным родословным знатных фамилий Европы, степень выраженности которых не изменялась на протяжении поколений.
Эти взгляды на наследственность были тесно связаны с представлениями о безграничной и ненаправленной изменчивости признаков и свойств живого, которая возникала как приспособительная реакция на воздействия внешних факторов. Подобные представления о характере наследственности и изменчивости, в общем, разделял и Ч. Дарвин, что нашло отражение в его концепции пангенеза.
Основы генетики, науки о наследственности, были заложены исследованиями каноника монастыря г. Брно, Чехия, Грегора Менделя. Он впервые экспериментально доказал дискретность наследования признаков и их независимое распределение среди потомков при возвратных скрещиваниях в последующих поколениях. Его работы опередили время и были признаны лишь в начале 20 века, спустя 25 лет после опубликования.
В этот период были сформулированы представления о мутациях как дискретных наследуемых изменениях свойств организмов (Г. де Фриз), представления о генах как единицах наследственности (В. Иогансен) без указания на их локализацию в клетке. Было введено также понятие " генотип " для обозначения наследственной конституции гамет и зигот в противоположность фенотипу – совокупности внешних, доступных непосредственному наблюдению признаков и свойств живых организмов.
В дальнейшем работами американских генетиков школы Моргана было установлено линейное расположение генов как локусов хромосомы в хромосомах ядра, явление рекомбинации генов между гомологичными хромосомами в результате генетического кроссинговера в процессе мейоза. Эти и другие работы послужили основой для формирования хромосомной теории наследственности, согласно которой материальными носителями свойств и признаков организмов служат дискретные элементы (локусы) хромосом.
До 50-х годов ХХ века молекулярная природа генов оставалась неизвестной. И лишь в 1953 г. была расшифрована структура ДНК и высказана гипотеза о её роли хранителя наследственной информации, принципах её кодирования и реализации при синтезе белка. Была сформулирована «центральная догма» генетики, воплотившая в молекулярной форме положение Вейсмана о невозможности передачи информации от зародышевой плазмы соматическим клеткам. Согласно ей наследственная информация может быть передана только в направлении ДНК→РНК→белок. На протяжении двух десятилетий были расшифрованы молекулярные механизмы передачи и реализации генетической информации в клетке, в результате хромосомная теория наследственности получила молекулярное обоснование. Считались доказанными: неизменное положение генов в хромосоме, однозначена связь гена и кодируемого им продукта (один ген – один фермент), функциональная активность всей ДНК в хромосомах ядра, зависимость сложности и уровня организации объектов от количества ДНК. Основные положения генетики этого периода отражены в таблице 4.1.
Таблица 4.1. Изменение представлений о структуре и функции генетического материала (Голубовский, 2000)
| Классическая генетика до начала 70-х годов | Подвижная генетика c 80-х годов |
| ДНК хранитель наследственной информации; структура ДНК код, все её изменения функционально важны. Чем больше ДНК в геноме, тем больше генов. Многообразие форм в природе есть отображение многообразия ДНК. | В хромосомах эукариот есть разные, заведомо неинформационные, "бессмысленные" фракции ДНК, состоящие из последовательностей, повторенных многие сотни, тысячи или миллионы раз. Близкие виды могут сильно отличаться по составу и количеству ДНК, имея одно и то же число генов. |
| Поток наследственной информации однонаправлен: ДНК→РНК→белок ("центральная догма" молекулярной биологии). | Возможен и обратный поток информации от РНК к ДНК с помощью фермента ревертазы; РНК способна быть ферментом; белки могут влиять на вторичную структуру ДНК, наследуемую в ряду поколений, возможен нематричный синтез ДНК. |
| Последовательности оснований в ДНК гена и матричной РНК строго соответствуют последовательностям аминокислот в белках (принцип "колинеарности") и физический размер гена соответствует размеру кодируемого им продукта. | У эукариот ген мозаичен; он состоит из кодирующих сегментов (экзонов) и некодирующих (интронов), которые вырезаются из матричной РНК. Между размером гена в хромосоме и размером кодируемого полипептида нет определенного соответствия. |
| Ген занимает определенное место в хромосоме (локус) и находится в одной или строго определенном числе копий у всех особей вида. | В геномах эукариот есть серия мобильных генов, число и топография которых варьирует от индивида к индивиду; гены могут амплифицироваться и менять число своих копий. |
| Репликация ДНК происходит только в ядре клетки. | В цитоплазме кроме автономно реплицирующихся ДНК митохондрий и пластид есть разные ДНК- и РНК-носители. |
| Некоторые фаги бактерий способны встраиваться в хромосому и существовать в ней в форме профага, а также переносить гены из одной бактерии в другую (трансдукция). | У всех эукариот происходит регулярное встраивание в хромосому хозяина последовательностей РНК и ДНК-содержащих вирусов. Вирусы - универсальный переносчик генов в биоценозе. |
| Виды - репродуктивно и генетически замкнутые системы. Симбиоз, подобный лишайникам, редкое, исключительное явление. | Наследственные системы эукариот полигеномны. Симбиоз генетических элементов разного происхождения привел к возникновению эукариот и регулярно происходит в природе. Гипотеза о потенциальном единстве генофонда всех видов в биоценозе. |
Исходя из этих положений, можно было предположить, что количество ДНК наиболее сходно у организмов, находящихся на одном уровне организации, и близкородственные виды обладают наибольшим сходством по количеству ДНК.
Однако оказалось, что при сходном количестве активных генов они могут значительно различаться по величине ДНК. Так, у некоторых рыб и амфибий количество ДНК в ядре в 25 раз превышает эту величину у любого вида млекопитающих, а у близких видов количество ДНК может различаться в несколько раз, в частности, вся ДНК генома риса может уместиться в одной из 42 хромосом мягкой пшеницы. Возник так называемый С-парадокс, который подрывал основы целого ряда устоявшихся представлений.
Современные представления о структуре генома
Термин «геном» введён в генетический обиход позднее понятия «генотип». В настоящее время они почти совпадают. Геном определяется как совокупность ядерных и цитоплазматических ДНК и РНК носителей с локализованными в них генетическими элементами, включая определенные функциональные (эпигенетические) связи между этими элементами. Таким образом, под геномом подразумевается вся наследственная система клетки.
Начиная с 70-х годов, стало появляться всё больше исследований, на основе которых становилось ясно, что механизмы хранения и передачи генетической информации значительно сложнее, чем это описывалось в рамках классической генетики 50-х – 60-х годов.
Было открыто явление обратной транскрипции – передачи генетической информации от РНК на ДНК, в частности, у РНК-содержащих вирусов (ретровирусы), которые при внедрении в клетку-хозяина обладают способностью синтезировать на своей РНК молекулы ДНК, которые затем встраиваются в ДНК клетки.
Был обнаружен нематричный синтез ДНК. В клетках иммунной системы (В-лимфоциты) при синтезе гена, кодирующего антитело, происходит связка фрагментов прежних генов, причем в точке их соединения в текст ДНК встраивается короткий фрагмент, нигде не хранимый, а синтезируемый и встраиваемый только ферментами. Важно то, что данная генетическая информация берётся не из генетического текста, а из функционального состояния ферментативной системы. В этом смысле она перенесена с белков на ДНК и означает переход: белок → ДНК.
Был открыт процесс, названный редактированием РНК. В ряде случаев информация, скопированная с ДНК на матрицу РНК, не соответствует структуре конечного продукта – белка. Однако перед его синтезом на матрице информационной РНК в рибосоме молекула РНК подвергается изменениям с помощью специальных ферментов, которые модифицируют её так, что она начинает кодировать синтез «правильного белка».
В ядре и цитоплазме клеток обнаружилось множество разновидностей внехромосомной ДНК. Ранее, при рассмотрении происхождения эвкариот, отмечалось наличие в митохондриях и хлоропластах собственной ДНК. Однако в ядре и цитоплазме клеток имеются и другие формы нехромосомной ДНК в виде кольцевых ДНК, плазмид, В-хромосом, вирусной ДНК. У прокариот также были найдены различные внехромосомные элементы: плазмиды, обладающие способностью к самостоятельной репликации, то есть к размножению в цитоплазме, а также бактериофаги, имеющие самостоятельную белковую оболочку. Эти элементы обладают способностью перехода из одной категории в другую, а также могут встраиваться в хромосомную ДНК.
Строение генов эвкариот оказалось отличным от генов прокариот. Они включают фрагменты двух типов: экзоны, принимающие участие в синтезе конкретного продукта, и интроны, никакого отношения к этому не имеющие. Количество последних могло превышать число активных фрагментов, они могут принимать участие в работе других генов. Следствием такой структуры генов эвкариот явилось значительное усложнение процесса транскрипции – сборки молекулы информационной РНК (иРНК), на которой происходит синтез белка в рибосоме, на матрице ДНК хромосомы. Сначала РНК включает в состав и копии интронов. Однако пред выходом РНК из ядра в цитоплазму особые ферменты – рестриктазы вырезают интроны, а экзоны сшиваются специальными ферментами – реализуется процедура сплайсинга. И только после него состоящая из одних экзонов иРНК переносится к рибосомам.
Одно из возможных преимуществ такой структуры эвкариотических генов заключается в следующем:
· блоки экзонов могут перемешиваться между собой и, в результате, гораздо быстрей, чем путём замены оснований, возникают новые генные конструкции;
· новые генные конструкции могут возникать и апробироваться под покровом старого гена, если интроны, в силу неточности вырезания (сплайсинга), будут иногда считываться вместе с экзонами. Такого рода явление получило название альтернативный сплайсинг. Следовательно, с одного участка ДНК (гена) могут считываться две или более РНК-матриц, и тогда образуется несколько вариантов белков. Альтернативный сплайсинг обнаружен у большинства генов. Он позволяет с одного гена получать разные продукты на разных этапах онтогенеза и в разных тканях
Было установлено, что гены, кодирующие синтез определённых продуктов - структурные гены – составляют незначительную часть ДНК хромосом (несколько процентов). Однако с ними функционально связаны гораздо более многочисленные гены-регуляторы, обеспечивающие изменение активности структурных генов, либо усиливающие, либо ослабляющие скорость синтеза конкретного продукта. С одним структурным геном может быть связано до нескольких десятков таких генов-регуляторов.
Функционально активная ДНК хромосом составляет меньшую часть всей ДНК. Значительная её доля, до 80–90% - неактивная (не транскрибируемая) ДНК, состоящая из коротких последовательностей нуклеотидов, повторяющихся сотни тысяч, иногда миллионы раз. Чаще всего эти участки образуют концевые отделы хромосом или отделы, примыкающие к центромерам, к которым прикрепляются нити веретена при делении клетки. Эти отделы микроскопически представлены облигатным хроматином, не изменяющимся при изменении функционального состояния клетки и представленным максимально конденсированными участками ДНК.
Длина последовательностей нуклеотидов и количество их повторов может быть иной, среди таких участков могут встречаться и функционально активные мультигенные семейства.
Другой особенностью генома, обнаруженной в последние десятилетия, является совокупность явлений, определённая как непостоянство генома. Сюда относится, прежде всего открытие ещё в 50-е годы прошлого века мобильных генетических элементов (МГЭ) у кукурузы. Это открытие получило признание лишь спустя 25 лет после их обнаружения у многих организмов самого различного типа. Оказалось, что некоторые генетические структуры способны менять своё положение, перемещаясь по хромосоме и встраиваясь в её новые участки.
У эвкариот МГЭ представлены транспозонами. Они обнаружены у кукурузы, дрозофилы (12 –25% ДНК хромосом), а также в виде элемента, широко распространённого в природе от нематод до человека. Спонтанные мутации у дрозофилы в 70% случаев связаны с вставками (инсерциями) в основном в виде МГЭ. У эвкариот вставки могут реализоваться и вирусоподобными ретротранспозонами, которые с помощью гена, кодирующего фермент ревертазу, строят на своей РНК ДНКовую копию, которая затем встраивается в хромосому.
Эти вставки (инсерции) могут вызывать разрывы хромосом, усиление мутационной активности в окрестностях новой точки локализации МГЭ, причем мутации носят нестабильный характер и могут возвращаться к норме. Важно отметить, что в этом случае они обычно носят массовый и определённый характер, и не похожи на случайные и единичные мутации классической генетики.
Следовательно, многие участки хромосомной ДНК содержат фрагменты, имеющие вирусное происхождение, а также представлены материалом плазмид и других нехромосомных разновидностей ДНК, встроенных в хромосомы.
Высказано предположение, что МГЭ играют роль своего рода рецепторов внешних стрессирующих сигналов, инициирующих вспышки транспозиционной изменчивости в критические периоды эволюции популяций, что может приводить к быстрому преобразованию гомеостатической видовой нормы и, возможно, — к видообразованию.
Основные изменения во взглядах на структуру генома на протяжении последних десятилетий ХХ века отражены в таблице 4.1.
Рассмотренные данные позволяют по-новому представить структуру генома и механизмы взаимодействия между его составными частями. Наиболее полно это представлено в концепции, предложенной М.Д.Голубовским. Согласно этой концепции, компоненты генома состоят из двух частей: облигатной компоненты (ОК) и факультативной компоненты (ФК). Состав и взаимосвязи между ними и внешней средой представлены на рис. 4.1.
ОК представлена генами хромосом ядра и органелл клетки – объектов исследований классической генетики. Состав ФК более разнообразен как по происхождению, так и по локализации в клетке. Важно, что между ОК и ФК возможен обмен элементами, причём переход ФК→ОК наблюдается чаще и происходит в виде инсерционных мутаций (вставок). Переход ОК→ФК – событие более редкое и является следствием амплификации (создания копий) генов хромосом и их обособления от хромосом.
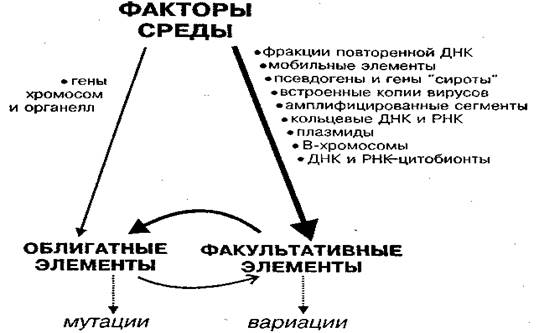
Рис.4.1. Пути возникновения наследственных изменений в природе в системе СРЕДА — ФАКУЛЬТАТИВНЫЕ — ОБЛИГАТНЫЕ ЭЛЕМЕНТЫ (Голубовский, 2000)
Факультативный компонент (ФК) генотипа образуют последовательности ДНК, количество и расположение которых могут свободно варьировать в разных клетках и у разных особей (вплоть до их полного отсутствия). Сюда входят также внутриклеточные, способные к автономной или полуавтономной репликации РНК-носители (плазмиды, вирусы). Важная особенность ФК по сравнению с ОК - неравномерность в распределении этих элементов между дочерними клетками при делении (например, в случае В-хромосом). Существуют как внутриядерные, так и цитоплазматические факультативные элементы. В ядре факультативные элементы расположены в хромосомах или не связаны с ними.
Среди хромосомных факультативных элементов можно выделить:
- фракции ДНК, которые расположены блоками и повторены сотни тысяч или миллионы раз. Они, как правило, не способны к транскрипции. Эти фракции называют сателлитными (стДНК), если они резко отличаются по составу нуклеотидов от остальной ДНК генома. Доля стДНК у разных видов составляет от 1 до 80% генома
- умеренно повторяющиеся последовательности (от 10 до 105 раз), составляющие 10–30% всей ДНК генома эукариот. Среди них есть элементы ОК в виде семейств повторенных жизненно важных генов, кодирующих белки рибосом, хромосомные белки - гистоны, транспортные РНК и т. д. Но основу умеренных повторов составляют элементы ФК, прежде всего, разного рода семейства рассеянных по геному мобильных генетических элементов. В геноме человека доля умеренных повторов составляет около 42%.
- рассеянные по геному осколки генов ОК — их копии, лишённые интронов, так называемые псевдогены, не способные к транскрипции, а также гены–сироты, одиночные копии семейств тандемно повторенных генов;
- эндогенные вирусы, частично или полностью интегрированные в разные участки хромосом хозяина; они встречаются у всех изученных видов эукариот).
В клеточном ядре обнаруживаются разного рода ДНК и РНК-носители, которые образуют дополнительные фракции к основному набору хромосом. Сюда могут входить:
- мелкие хромосомные образования, двойные микрохромосомы и т. д.;
- внутриядерные симбионты в виде вирусов или бактерий;
- добавочные или В-хромосомы. Они находятся в особом гетерохроматиновом состоянии, число их варьирует у разных особей одного вида и в разных тканях одного организма. В-хромосомы найдены более чем у 700 видов растений, у сотен видов беспозвоночных и около 20 видов позвоночных.
Взаимосвязи и взаимодействие между этими двумя компонентами генома будут рассмотрены при описании форм изменчивости.

