 2015-10-16
2015-10-16 2628
2628Любая молекула может пройти через липидный бислой, однако скорость пассивной диффузии веществ, т.е. перехода вещества из области| с большей концентрацией в область с меньшей может сильно отличаться. Для некоторых молекул это занимает столь длительное время, что можно говорить об их практической непроницаемости для липидного бислоя мембраны. Скорость диффузии веществ через мембрану зависит главным образом от размера молекул и их относительной растворимости в жирах.
Легче всего проходят простой диффузией через липидную мембрану малые неполярные молекулы, такие как О2, стероиды, тиреоидные гормоны, а также жирные кислоты. Малые полярные незаряженные молекулы — СО2, NН3, Н2О, этанол, мочевина — также диффундируют с достаточно большой скоростью. Диффузия глицерола идёт значительно медленнее, а глюкоза практически не способна самостоятельно пройти через мембрану. Для всех заряженных молекул, независимо от размера, липидная мембрана непроницаема.
Транспорт таких молекул возможен благодаря наличию в мембранах либо белков, формирующих в липидном слое каналы (поры), заполненные водой, через которые могут проходить вещества определённого размера простой диффузией, либо специфических белков-переносчиков, которые избирательно взаимодействуя с определёнными лигандами, облегчают их перенос через мембрану (облегчённая диффузия).
Кроме пассивного транспорта веществ, в клетках есть белки, активно перекачивающие определённые растворённые в воде вещества против их градиента, т.е. из меньшей концентрации в область большей. Этот процесс, называемый активным транспортом, осуществляется всегда с помощью белков-переносчиков и происходит с затратой энергии.
строение и функционирование БЕЛКОВЫХ КАНАЛОВ
Каналы в мембране формируются интегральными белками, которые «прерывают» липидный бислой, образуя пору, заполненную водой. Стенки канала «выстилаются» радикалами аминокислот этих белков.
Если каналы различают вещества только по размеру и пропускают все молекулы меньше определённой величины, по градиенту концентрации, т.е. служат фильтрами, то их называют «неселективные каналы», или «поры». Такие поры есть в наружной мембране митохондрий, где молекулы белка-порина образуют широкие гидрофильные каналы. Через них могут проходить все молекулы с молекулярной массой 10 кД и меньше, в том числе и небольшие белки.
Селективные каналы, как правило, участвуют в переносе определённых ионов. Ионная селективность (избирательность) каналов определяется их диаметром и строением внутренней поверхности канала. Например, катионселективные каналы пропускают только катионы, так как содержат много отрицательно заряженных аминокислотных остатков.
Открытие или закрытие селективных каналов регулируется либо изменением концентрации специфических регуляторов, таких как медиаторы, гормоны, циклические нуклеотиды, N0, G-белки, либо изменением трансмембранного электрохимического потенциала. Воздействие регуляторного фактора вызывает конформационные изменения каналообразующих белков, канал открывается и ионы проходят по градиенту концентрации. Транспорт веществ через каналы не приводит к конформационным изменениям белков и зависит только от разности концентраций веществ по обе стороны мембраны. Поэтому скорость транспорта веществ через такие каналы может достигать 106-108 ионов в секунду.
облегчённая диффузия веществ
В мембранах клеток существуют белки-транслоказы. Взаимодействуя со специфическим лигандом, они обеспечивают его диффузию (транспорт из области большей концентрации в область меньшей) через мембрану. В отличие от белковых каналов, транслоказы в процессе взаимодействия с лигандом и переноса его через мембрану претерпевают конформационные изменения. Кинетически перенос веществ облегчённой диффузией напоминает ферментативную реакцию. Для транслоказ существует насыщающая концентрация лиганда, при которой все центры связывания белка с лигандом заняты, и белки работают с максимальной скоростью Vmax. Поэтому скорость транспорта веществ облегчённой диффузией зависит не только от градиента концентраций переносимого лиганда, но и от количества белков-переносчиков в мембране.
Существуют транслоказы, переносящие только одно растворимое в воде вещество с одной стороны мембраны на другую. Такой простой транспорт называют «пассивный унипорт». Примером унипорта может служить функционирование ГЛЮТ-1 — транслоказы, переносящей глюкозу через мембрану эритроцита.
Некоторые транслоказы могут переносить два разных вещества по градиенту концентраций в одном направлении — пассивный симпорт, или в противоположных направлениях — пассивный антипорт.
Примером транслоказы, работающей по механизму пассивного антипорта, может служить анионный переносчик мембраны эритроцитов.
Внутренняя митохондриальная мембрана содержит много транслоказ, осуществляющих пассивный антипорт. В процессе такого переноса происходит эквивалентный обмен ионами, но не всегда эквивалентный обмен по заряду.
строение и функционирование БЕЛКОВ-ПЕРЕНОСЧИКОВ, ОСУЩЕСТВЛЯЮЩИХ АКТИВНЫЙ ТРАНСПОРТ
Перенос некоторых лигандов (ионов, глюкозы, аминокислот) через мембраны происходит против градиента концентрации и сопряжён с затратой энергии (активный транспорт). Перенос лигандовчерез мембрану, связанный с затратой энерги АТФ, называют «первично-активный транспорт»
I. Первично-активный транспорт
Перенос некоторых неорганических ионов идёт против градиента концентрации при участии транспортных АТФ-аз (ионных насосов) Все ионные насосы одновременно служат ферментами, способными к аутофосфорилированию и аутодефосфорилирова-нию. АТФ-азы различаются по ионной специфичности, количеству переносимых ионов, направлению транспорта. В результате функционирования АТФ-азы переносимые ионы накапливаются с одной стороны мембраны. Наиболее распространены в плазматической мембране клеток человека Nа+,К+-АТФ-аза, Са2+-АТФ-аза и Н+,К+,-АТФ-аза слизистой оболочки желудка.
Nа+,К+-АТФ-аза
Этот фермент-переносчик катализирует АТФ-зависимый транспорт ионов Nа + и К+ через плазматическую мембрану. Na+,К+-АТФ-аза состоит из субъединиц α и β; α — каталитическая большая субъединица, а β — малая субъединица (гликопротеин). Активная форма транслолоказы тетрамер (αβ)2.
Nа+,К+-АТФ-аза отвечает за поддержание высокой концентрации К+ в клетке и низкой концентрации Na+. Так как Na+,К+-АТФ-аза выкачивает три положительно заряженных иона, а закачивает два, то на мембране возникает электрический потенциал с отрицательным значением на внутренней части клетки по отношению к её наружной поверхности.
Са2+-АТФ-аза
В цитозоле «покоящихся» клеток концентрация Са2+ составляет ~10-7 моль/л, тогда как вне клетки она равна ~2-10-3 моль/л. Поддерживает такую разницу в концентрации система активного транспорта ионов кальция; её основные компоненты — кальциевые насосы — Са2+-АТФ-азы и Na+,Са2+ -обменники.
Са2+-АТФ-аза локализована не только в плазматической мембране, но и в мембране ЭР. Фермент состоит из десяти трансмембранных доменов, пронизывающих клеточную мембрану. Между вторым и третьим доменами находятся несколько остатков аспарагиновой кислоты, участвующих в связывании кальция. Область между четвёртым и пятым доменами имеет центр для присоединения АТФ и аутофосфорилирования по остатку аспарагиновой кислоты. Са2+-АТФ-азы плазматических мембран некоторых клеток регулируются белком кальмодулином. Каждая из Са2+-АТФ-аз плазматической мембраны и ЭР представлена несколькими изоформами.
Работа Са2+-АТФ-азы цитоплазматической мембраны по стадиям представлена на рис.
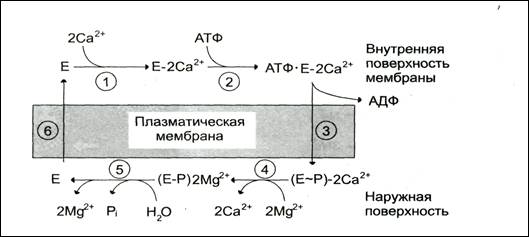
Последовательность событий в процессе работы Са2+-АТФ-азы
1 – связывание двух ионов кальция участком АТФ-азы, обращенной в цитозоль; 2 – изменение заяда и конформации фермента (АТФ-азы), вызванное присоединением двух иоов кальция – приводит к повышению сродства к АТФ и активации аутофосфорилирования; 3- аутофосфорилирование – сопровождается конформационными изменениями, АТФ-аза закрывается с внутренней стороны мембраны и открывается с наружной; 4 – происходит снижение сродства центров связывания к ионам кальция и они отделяются от АТФ-азы; 5 – аутодефосфорилирование, активируется ионами магния, в результате Са-АТФ-за теряет фосфорный остаток и два иона магния; 6 – АТФ-аза возвращается в исходное состояние.
Нарушение активности Са2+-АТФ-азы при патологии. Одна из причин нарушения работы Са2+-АТФ-азы — активация перекисного окисления липидов (ПОЛ) мембран. Окислению подвергаются как ацильные остатки жирных кислот в составе фосфолипидов, так и SН-группы в активном центре фермента. Нарушение структуры липидного окружения и структуры активного центра приводит к изменению конформации АТФ-азы, потере сродства к ионам кальция и способности к аутофосфорилированию. АТФ-аза перестаёт выкачивать ионы кальция из цитозоля клетки, повышается концентрация внутриклеточного кальция; Са2+ усиливает мышечное сокращение, возрастает тонус мышечной стенки, что приводит к повышению АД. Нарушение функционирования Са2+-АТФ-азы играет свою роль в развитии атеросклероза, рака, иммунных патологий.

