 2015-02-27
2015-02-27 8278
8278Для всех микроорганизмов, входящих в царство, характерен прокариотический тип организации клетки, что определяется особенностями их ультраструктуры, а также строения и функций ряда макромолекул. Из всех известных клеток прокариотическая самая простая и, вероятно, первая клетка, возникшая около 3,6 млрд. лет назад.
В настоящее время предполагают, что в какой-то момент времени эволюция клеток пошла по двум самостоятельным направлениям. Появились две группы организмов —прокариоты, у которых ядерный материал не был ограничен оболочкой, и эукариоты, имеющие оформленное ядро с ядерной оболочкой.
Основные отличия прокариотов от эукариотов состоят в следующем:
- в клетках прокариотов отсутствуют компартменты, или органеллы, ограниченные от цитоплазмы специализированными внутриклеточными липопротеидными мембранами: эндоплазматическая сеть (ретикулюм), митохондрии, аппарат Гольджи, лизосомы, хлоропласты;
- ядерная структура прокариотов, называемая нуклеоидом, не имеет ядерной оболочки с поровым комплексом и представлена макромолекулой ДНК с белками (без гистонов);
- геном прокариотической клетки организован в одну кольцевую хромосому, которая представляет собой единый репликон и не делится митозом;
- дополнительные репликоны могут быть представлены кольцевыми молекулами плазмидных ДНК;
- прокариотическая клетка содержит только один тип рибосом с константой седиментации 708, причем часть рибосом ассоциирована с цитоплазматической мембраной, что никогда не наблюдается у эукариотов;
- клеточная стенка прокариотов содержит характерный только для бактерий биогетерополимер — пептидогликан.
Некоторые прокариоты обладают структурами, отсутствующими у эукариотов:
- подвижные бактерии имеют особые бактериальные жгутики из белков-флагеллинов;
- спорообразующие формы бактерий в неблагоприятных условиях превращаются в уникальные по степени устойчивости типы покоящихся клеток — бактериальные споры;
- клетки прокариотов очень малы; диаметр большинства клеток бактерий не превышает 1 мкм, однако длина может быть значительной, например, у некоторых спирохет — до 500 мкм. Малые размеры прокариотов связаны, как полагают, с отсутствием в их ультраструктуре специализированных мембранных систем, что затрудняет координацию внутриклеточных процессов пропорционально увеличению размеров клетки.
Клеточное строение четко отделяет прокариотов от вирусов. Подчеркивая примитивность организации бактериальных клеток, необходимо однако отметить, что они эволюционировали в своем направлении в течение гораздо большего времени, чем эукарио-тические, и, хотя эволюционные возможности прокариотической клетки, по всей видимости, ограничены, в процессе эволюции происходили изменения их клеточной организации, что привело постепенно к ее усложнению.
По целому ряду признаков бактерии имеют принципиальные различия с эукариотами, и знание особенностей их устройства и функционирования позволяет понять возможность избирательного антимикробного действия химиотерапевтических препаратов. Применение электронной микроскопии и тонких цитохимических исследований позволило изучить их ультраструктуру (рис. 1). Обязательными компонентами бактериальной клетки являются цитоплазматическая мембрана, окружающая цитоплазму, в которой содержатся рибосомы и нуклеоид. Клетки всех бактерий, за исключением L-форм и микоплазм, имеют клеточную стенку. Другие структуры являются дополнительными и определяют морфологические и функциональные особенности различных видов: капсулы, жгутики, пили, споры, включения.
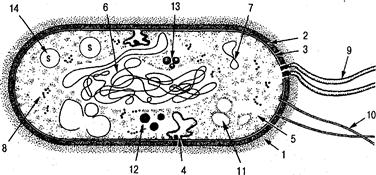
Рис. 1. Схема строения прокариотической клетки:
/ — капсула; 2 — клеточная стенка; 3 — цитоплазматическая мембрана; 4 — мезосомы; 5 — цитоплазма; 6 — нуклеоид; 7 — плазмида;
8 — рибосомы и полисомы; 9 — жгутики; 10 — пили; 11 — гранулы гликогена; 12 — липидные капельки; 13 — гранулы волютина; 14 — включения серы
Поверхностные структуры. Капсула — это наружный, самый верхний слизистый слой клетки различной толщины фибриллярной или глобулярной структуры. Она имеет полисахаридную, му-кополисахаридную или полипептидную природу и содержит до 98 % воды. В зависимости от толщины различают микрокапсулу (толщиной менее 0,2 мкм) и макрокапсулу. Капсула не является обязательным структурным элементом клетки. Биологический смысл капсулообразования определяется целым рядом функций, среди которых: защита от фагоцитов и вирусов, токсинов и радиации; иммунологическая мимикрия у патогенных бактерий; сохранение влаги в условиях пониженной влажности; прикрепление клетки к плотной поверхности.
Пили (фимбрии, ворсинки, реснички) — это прямые цилиндрические образования белковой природы длиной 0,3—10 мкм, диаметром до 10 нм, равномерно покрывающие поверхность клетки (до нескольких сотен на клетку), не выполняющие локомоторную функцию.
Различают пили общего типа, способствующие прикреплению бактериальной клетки к субстрату, клеткам человека (явление адгезии микроорганизмов) и пили половые, участвующие в передаче генетического материала от клетки-донора к клетке-реципиенту в процессе конъюгации, а также обуславливающие адсорбцию специфических бактериофагов на клетках.
Жгутики — органы движения бактерий в виде спирально изогнутых цилиндрических образований белковой природы (белки-флагеллины) на поверхности клетки длиной 3—12 мкм и толщиной 10—30 нм, прикрепленные базальным телом (системой дисков) к цитоплазматической мембране (см. вкл. I). Число и расположение жгутиков может быть различным и является видовым признаком (рис. 2). Различают монотрихи (бактерии с одним жгутиком на конце), амфитрихи (бактерии с жгутиками, расположенными по полюсам), лофотрихи (клетки с пучком жгутиков на одном конце) и перитрихи (с 2—30 жгутиками по всему телу клетки).
Пили и жгутики не являются обязательными органоидами бактериальной клетки.
Клеточная стенка — один из основных структурных элементов бактерии, выполняющий механическую защиту клетки. Кроме микоплазм и L-форм, клетки всех бактерий покрыты клеточной стенкой, толщина которой у разных видов колеблется в пределах 0,01—14 мкм. Она представляет собой плотную эластичную структу -
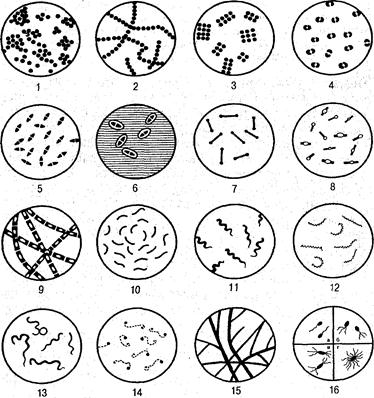
Рис. 2. Основные формы бактерий (по А. А. Воробьеву и соавт., 1994):
/ — стафилококки; 2 — стрептококки; 3 — сарцины; 4 — гонококки;
5— пневмококки; 6— капсулы пневмококков; 7— коринебактерии дифтерии; 8 — клостридии; 9 — бациллы; 10 — вибрионы; 11 — спириллы; 12 — трепонсмы; — боррелии; 14 — лептоспиры; 15— актиномицеты; 16 — расположение жгутиков: а — монотрихи; б — лофотрихи; в — амфитрихи; г — перитрихиру, которая окружает протопласт клетки и придает ей постоянную форму и жесткость. Клеточная стенка препятствует осмотическому набуханию и разрыву клеток, когда они попадают в гипотоническую среду. Вода, другие малые молекулы и разные ионы легко проникают через крошечные поры в клеточной стенке, но через них не проходят крупные молекулы белков и нуклеиновых кислот.
Основным химическим компонентом клеточной стенки является специфический гетерополимер — пептидогликан (муреин, мукопептид, глюкозаминопептид, гликопептид), состоящий из цепочек, в которых чередуются остатки N-ацетилглюкозамина и М-ацетилмурамовой кислоты, соединенные между собой β-1,4-гликозидными связями. Это резко отличает оболочечные структуры бактерий от эукариотических и создает «ахиллесову пяту» бактерий, используемую для антимикробной химиотерапии.
Организация цитоплазмы. Цитоплазматическая мембрана (ЦМ) Носится к числу обязательных клеточных структур, имеет толщину 7—13 нм и располагается непосредственно под клеточной стенкой, ограничивая протопласт клетки. По своему строению мембраны бактериальных, животных и растительных клеток очень сходны. В настоящее время большинством ученых принята жидкостно-мозаичная модель строения ЦМ. Согласно этой модели ЦМ состоит из двойного слоя (15—30 % молекул фосфолипидов и триглицеридов с направленными внутрь гидрофобными концами и гидрофильными «головками» наружу. В него мозаично погружены молекулы белка (50—70%). В мембране имеются также углеводы (2—5 %) и РНК. ЦМ представляет собой пластическое «текучее» образование, которое играет важнейшую роль обмене веществ, является полупроницаемой структурой, поддерживает осмотическое давление, контролирует как поступление веществ в клетку, так и выведение конечных метаболитов по системе субстрат специфичных пермеаз (ферментов-переносчиков, локализованных на мембране). С ЦМ связаны процессы дыхания, доставляющие клетке энергию, то есть те функции, за которые в эукариотической клетке ответственны мембраны митохондрий и хлоропластов.
Выделяют так называемые мезосомы — впячивания ЦМ — смешанные мембранные системы, образованные трубочками, пузырьками и ламеллами. Предполагается выполнение ими функций центра дыхательной активности бактерий, участие в делении клетки и расхождении дочерних хромосом после репликации.
Цитоплазма заполняет собой объем бактерии, ограниченный ЦМ. Это сложная коллоидная система, которая состоит из бельков, нуклеиновых кислот, углеводов, липидов, минеральных веществ и 70—80 % воды. Цитоплазма является местонахождением внутриклеточных органелл (нуклеоида, рибосом, различных включений) и участвует во внутриклеточном метаболизме. Характерными чертами организации цитоплазмы прокариотов по сравнению с животными и растительными клетками являются отсутствие эндоплазматического ретикулюма и высокая электронная плотность.
Нуклеоид — ядерный материал бактериальной клетки. Он представлен двойной нитью макромолекулы ДНК с молекулярной массой 2—310 в соединении с белками, среди которых отсутствуют характерные для эукариотов ядерные (гистоны и гистоноподобные) белки. В отличие от настоящего ядра эукариотических клеток нуклеоид не имеет ядерной перфорированной мембраны, не делится митозом и представляет собой в период деления одну кольцевую хромосому, кодирующую всю генетическую информацию.
Плазмиды — необязательные внутриклеточные структуры в виде внехромосомных кольцевых участков ДНК, способных к саморепликации. Обуславливают наследование дополнительных признаков: лекарственной устойчивости, токсигенности, бактериоциногенности и др.
Рибосомы — органеллы, в которых осуществляется синтез белка. Каждая рибосома имеет размеры 20х30х30 нм и константу седиментации 70S (так как при ультрацентрифугировании рибосомы оседают со скоростью около 70 единиц Сведенберга (S), в отличие от более крупных цитоплазматических рибосом эукариотов с константой седиментации 808). В свободном состоянии бактериальная рибосома находится в виде двух субъединиц — 30S и 50S, обе субъединицы содержат примерно по 40 % рибосомальной РНК и 60 % белка. Во время синтеза белка рибосомы с помощью информационной РНК образуют полисомы, обычно связанные с ЦМ. Бактерии могут содержать от 5000 до 50 000 рибосом, что зависит от возраста клетки и условий культивирования.
Знание различий между рибосомами бактерий и эукариотических клеток имеет важное значение для понимания механизмов антимикробного действия тех антибиотиков, которые подавляют синтез белка на бактериальных рибосомах и не затрагивают функции 80S рибосом.
Споры (эндоспоры) бактерий — покоящиеся формы некоторых видов грамположительных бактерий в неблагоприятных условиях внешней среды.
Спорообразование происходит в несколько стадий, при полном созревании споры вегетативная часть клетки лизируется и отмирает (см. вкл. I, II).
В процессе спорообразования (споруляции) можно выделить несколько основных этапов. Переходящая к спорообразованию клетка перестает расти; как правило, она содержит два и более нуклеоидов. На первом этапе часть клеточной ДНК локализуется в одном из полюсов клетки. Затем часть цитоплазмы с заключенной в ней хромосомой обособляется цитоплазматической мембраной, как бы врастающей в глубь клетки, при этом образуется проспорь, окруженная двойной мембранной оболочкой.
Затем между двумя мембранами идет формирование многослойной стенки и коры (кортекса) споры пептидогликановой природы. Снаружи мембран образуется также полипептидная оболочка и экзоспорий, окружающий спору в виде свободного чехла. Полностью сформировавшаяся бактериальная спора — это уплотненный участок клетки с нуклеоидом и рибосомами, ограниченный плотной многослойной оболочкой, пропитанной кальциевыми Солями дипиколиновой кислоты.
Спорообразование характерно для палочковидных бактерий — бацилл и клостридий (см. рис. 2). Различают центральное, терминальное и субтерминальное расположение спор в вегетативной части клетки, что является дифференциально-диагностическим признаком возбудителя.
В одной бактерии образуется одна спора, находящаяся в стадии покоя, при этом все процессы обмена веществ практически сведены к нулю, но сохраняется потенциальная жизнеспособность клетки. Поскольку увеличение числа микроорганизмов в этом процессе не происходит, спорообразование у бактерий не является способом размножения, а лишь приспособлением для выживания. Уникальные по степени своей устойчивости к физическим и химическим факторам бактериальные споры могут сохраняться до внешней среде без потери жизнеспособности длительное время (десятки лет), затрудняя борьбу со спороносными патогенными бактериями.
Внутриплазматические включения. Термином «включения» обозначают такие внутриклеточные структуры бактерий, которые, очевидно, не являются абсолютно необходимыми для их жизнедеятельности. Однако их природа и функции могут быть различны. В одних случаях включения являются продуктами обмена бактериальной клетки, в других — запасом питательных веществ.
Из резервных полисахаридов особенно распространены глюканы — гликоген, крахмал, гранулеза. Они выявляются в клетках бацилл, клостридий, энтеробактерий и др.
Запасные липиды представлены полиэфиром β-оксимасляной кислоты и восками. Воски, эфиры высокомолекулярных жирных кислот и спиртов характерны для микобактерий.
У коринебактерий резерв фосфора создается в виде зерен полифосфатов (волютина), имеющих диагностическое значение.

