 2015-02-24
2015-02-24 4289
4289Пигменты — важнейший компонент аппарата фотосинтеза. Изучение растительных пигментов резко ускорилось благодаря работам русского физиолога растений М. С. Цвета. Пытаясь найти способ разделения пигментов на индивидуальные вещества, Цвет в 1901 — 1903 гг. открыл принципиально новый метод, который он назвал адсорбционной хроматографией. Через колонку с сорбентом пропускается растворитель с растворенными веществами. Так как вещества различаются по степени адсорбции, они перемещаются по колонке с разной скоростью. В результате происходит разделение веществ.
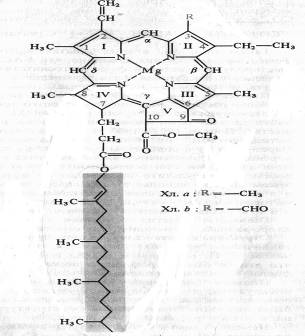
Польские биохимики М. Ненцкий и Л. Мархлевский (1897) обнаружили, что основу молекулы хлорофилла, как и гема гемоглобина, составляет порфириновое кольцо. Таким образом было показано принципиальное структурное сходство этих пигментов у растений и животных.
Немецкий химик Р. Вильштеттер в 1906—1914 гг. установил элементарный состав хлорофилла а — C55H72O5N4Mg и хлорофилла b — C55H70O6N4Mg. Хлорофилл — сложный эфир дикарбоновой кислоты хлорофиллина, у которой одна карбоксильная группа этерифицирована остатком метилового спирта, а другая остатком одноатомного непредельного спирта фитола.
|
|
|
Четыре пиррольных кольца (I — IV) соединены между собой метановыми мостиками, образуя порфириновое ядро. Наружные атомы углерода в тетрапиррольном цикле обозначены цифрами 1 — 10. Атомы азота пиррольных колец четырьмя координационными связями взаимодействуют с атомом магния. В структуре порфиринового ядра есть также циклопентановое кольцо (V), образованное остатком кетопропионовой кислоты и содержащее химически активную карбонильную группу у С9 и метилированную карбоксильную группу у С10. Структура, состоящая из тетрапиррольного и циклопентанового колец, получила название форбина. Боковая цепь IV пиррольного кольца включает в себя пропионовую кислоту, связанную сложноэфирной связью с полиизопреновым непредельным спиртом фитолом (С20Н39ОН). У 1, 3, 5 и 8-го углеродов пиррольных колец имеются метальные группы, у 2-го — винильная, у 4-го — этильная группа. Порфириновое кольцо представляет собой систему из девяти пар конъюгированных (сопряженных) чередующихся двойных и одинарных связей с 18 делокализованными p-электронами. Хлорофилл b отличается от хлорофилла а тем, что у 3-го углерода вместо метильной находится формильная (—СНО) группа. Структура хлорофилла, лишенная фитола, называется хлорофиллидом. При замещении атома магния протонами в молекуле хлорофиллов образуются соответствующие феофетины. Все фотосинтезирующие растения, включая все группы водорослей, а также цианобактерии, содержат хлорофиллы группы а. Хлорофилл b представлен у высших растений, у зеленых водорослей и эвгленовых. У бурых и диатомовых водорослей вместо хлорофилла b присутствует хлорофилл c, а у многих красных водорослей — хлорофилл d.
|
|
|
Биосинтез хлорофиллов. Первый этап биосинтеза хлорофиллов у растений — образование 5-аминолевулиновой кислоты (А Л К) из С5-дикарбоновых кислот. Показано, что глутаминовая кислота через 2-гидроксиглутаровую превращается в 4,5-диоксовалериановую, которая затем аминируется за счет аланина или других аминокислот (рис. 5.2). Реакция переаминирования катализируется АЛК-трансаминазой с участием пиридоксальфосфата в качестве кофермента. Для синтеза АЛК может использоваться и оскетоглутаровая (2-оксоглутаровая) кислота. Циклизация двух молекул АЛК приводит к образованию пиррольного соединения — порфобилиногена. Из четырех пиррольных колец формируется уропорфириноген, который превращается в протопорфирин IX. Дальнейший путь превращений протопорфирина может быть различным. С участием железа формируется гем, входящий в состав цитохромов, каталазы, пероксидазы и гемоглобина. Если молекулу протопорфирина включается магний, затем карбоксильная группа у С10 этерифицируется метильной группой 6-аденозил-b-метионина и замыкается циклопентановое кольцо (К), то образуется протохлорофиллид. Под действием света в течение нескольких секунд протохлорофиллид превращается в хлорофиллид а в результате гидрирования двойной связи у С7 — С8 в IV пиррольном ядре. У низших растений и некоторых голосеменных (у хвойных) хлорофиллид может образовываться в темноте. Хлорофиллид обладает теми же спектральными свойствами, что и хлорофилл. Последний этап в формировании молекулы хлорофилла а — этерификация фитолом, который, как и все полиизопреновые соединения, синтезируется из ацетил-KоА через мевалоновую кислоту. По данным А. А. Шлыка (1965), хлорофилл b может образовываться из вновь синтезированных молекул хлорофилла а.
Растворы хлорофиллов в полярных растворителях обладают яркой флуоресценцией (люминесценцией). В этиловом эфире у хлорофилла а наблюдается рубиново-красная флуоресценция с максимумом 668 нм, у хлорофилла b — 648 нм
Наиболее устойчивы те состояния атомов и молекул, в которых валентные электроны занимают самые низкие энергетические уровни и распределены по ним согласно принципу Паули (не более двух электронов с антипараллельными спинами на каждой орбитали). Такое состояние молекулы называют основным синглетным (S0) энергетическим состоянием. Суммарный спин (вектор магнитных моментов) всех электронов молекулы в этом случае равен нулю. Когда молекула поглощает квант света, то за счет этой энергии электрон переходит на более высокие вакантные орбитали. Если у возбужденного электрона сохраняется то же направление спина, то молекула находится в синглетном возбужденном состоянии (S*). Если при переходе на более высокую орбиталь спин электрона обращается, то такое возбуждение называется триплетным (T*). Прямой переход из основного состояния в триплетное — маловероятное событие, так как для этого необходимо обращение ориентации спина. Переходу молекулы из основного состояния в возбужденное, происходящему при поглощении кванта света, соответствует полоса в спектре поглощения.
Поглощение молекулой хлорофилла кванта красного света приводит к синглетному электронвозбужденному состоянию — S*1.
При поглощении кванта синего света с более высоким уровнем энергии электрон переходит на более высокую орбиту (S*2). Возбужденная молекула хлорофилла возвращается в основное состояние различными путями. Отдав часть энергии в виде теплоты (особенно с уровня S*1), молекула может излучить квант света с большей длиной волны (правило Стокса),что проявляется в виде флуоресценции. Хлорофилл а поглощает свет в красной и синей областях спектра, но флуоресцирует лишь в красной. Время жизни синглетного возбужденного состояния 10-13 10-9с.
|
|
|
Другой путь потери энергии состоит в переходе молекулы из синглетного возбужденного состояния в метастабильное триплетное (с обращением спина). Этот переход безизлучательный, т. е. энергия теряется в виде теплоты. Триплетное возбужденное состояние имеет гораздо большее время жизни (>10-4 с). Из триплетного состояния молекула может вернуться в основное, излучив еще более длинноволновый (чём в случае флуоресценции) квант света. Это обычно более слабое свечение и есть фосфоресценция.
Энергия возбужденного состояния может быть использована на фотохимические реакции. В этом случае флуоресценции и фосфоресценции хлорофилла не наблюдается.
Молекула хлорофилла способна выполнять три важнейшие функции: 1) избирательно поглощать энергию света, 2) запасать ее в виде энергии электронного возбуждения, 3) фотохимически преобразовывать энергию возбужденного состояния в химическую энергию первичных фотовосстановленных и фотоокисленных соединений.

