 2015-08-12
2015-08-12 2849
2849Людське око дозволяє бачити предмети розміром не менше 0.1 мм., що багато разів більше діаметра більшості клітин. Тому відкриття клітини і розвиток клітинної теорії тісно пов’язані з розвитком оптики і створення мікроскопа.
Перший мікроскоп, що складався з двох чи більше лінз, було винайдено голландськими механіками братами Янсен у 1600 році. Згодом його вдосконалив англійський фізик Роберт Гук. Виступаючи в Лондоні на зборах Королівського товариства він продемонстрував можливості свого приладу, популярність мікроскопа зросла.
У 1672 році італійський біолог і лікар Марчелло Мальпігі у своїй книзі «Анатомія рослин» докладно описує мікроскопічні структури рослинних тканин. В цей же час нідерландський натураліст Антоні Ван Левенгук, використовуючи мікроскоп, спостерігав і змалював сперматозоїди, еритроцити, найпростіших та інше.
За десятки років, що пройшли після відкриття Р.Гука, величезне число дослідників використовувало мікроскопи, накопичилась велика кількість повідомлень, описів і малюнків різних типів клітин, тканин і мікроорганізмів. Однак усі вченні того часу головною частиною клітин вважали їх стінки, не приділяючи належної уваги внутрішньоклітинним структурам. Так продовжувалося аж до ХІХ століття.
Так, у 1835 р. чеський натураліст Ян Пуркіньє вперше описує ядро тваринної клітини, вводить термін «протоплазма» і стверджує, що саме вона, а не клітина стінка, є живою речовиною. В 1826 р. російський вчений Карл Бер відкриває яйцеклітину і вивчає ембріогенез, що затвердило його засновника ембріології як науки. В 1830 р. англійський ботанік Роберт Браун описує ядро рослинної клітини, вказуючи на те що ця структура є обов’язковим компонентом усіх клітин. У 1846р. німецький ботанік Гуго фон Моль дає класифікацію тканинам рослин, та розподіляє поняття «протоплазма» і «клітинний сік».
Після визначення клітинної теорії вона почала інтенсивно розвиватися й уточнюватися. Наприкінці ХІХ століття німецький медик і біолог Рудольф Вірхов довів, що клітини утворюються з інших клітин шляхом їх поділу. За короткий час (1883-1898рр.) відкрито пластиди рослинних клітин, більш глибоко вивчено клітинний поділ і описані хромосоми, виявлені мітохондрії й апарат Гольджі.
У 30-40 року ХХ сторіччя з винаходом електронного мікроскопа почалася нова ера в розвитку біології. Стало можливим вивчити ті деталі клітин, про які раніше людина не мала уявлення. З цього часу в цитології і фізіології рослин наступив різкий перелом - замість роздільного вивчення структури клітин і їхніх функцій вони стали розглядатися в органічній єдності, що дозволило розкрити найважливіші фізіологічні процеси, які в них протікають.
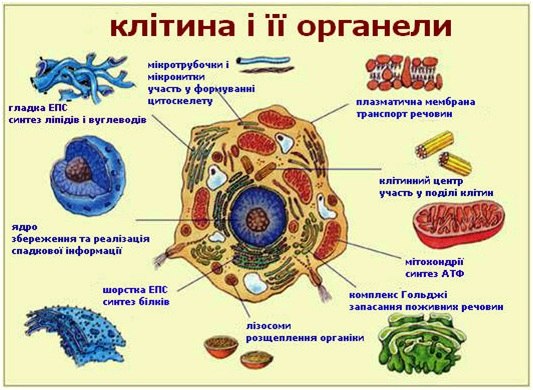
Клітина – це одиниця життя на нашій планеті, поза клітини життю немає. Саме тому всі особливості життєдіяльності організмів визначаються, виходячи з характеристик клітини, які визначають органели клітини і їх функції.
З безлічі властивостей клітини найбільш важливими вважають: строго певну будову, здатність до метаболізму – виробництву обміну речовин і енергії, здатність реагувати на зовнішні подразники: об’єкти та явища.
Крім того, функції органоїдів клітини визначають такі властивості, як здатність до зростання і розвитку, а також розмноження і відновлення – регенерацію.
Подібно до того, як організм тварини або рослини складається з окремих органів і їх систем, клітина складається з органоїдів.
Розглядаючи органели клітини і їх функції, важливо відзначити зовнішня будова клітини. Зовні «одиниця життя» покрита мембраною, яка служить розмежувальних бар’єром, відокремлюючи зовнішнє середовище від внутрішнього вмісту клітини.
При цьому мембрана виконує захисну і розмежувальну функції, а також бере подразники зовнішнього середовища (рецепторна функція) і здійснює транспорт речовин.
Органели - постійні внутрішньоклітинні структури, що мають певну будову і виконують відповідні функції.
Органели діляться на дві групи: мембранні та немембранні.
До не мембранні органел клітини відносяться центріолі, мікротрубочки, філаменти, рибосоми і полісоми.
Історично, органели були виявлені за допомогою різноманітних форм мікроскопії або завдяки клітинному фракціонуванню.
Рецептори та інші дрібні, молекулярного рівня, структури, органелами не називають. Кордон між молекулами і органелами дуже нечіткий. Так, рибосоми, які зазвичай однозначно відносять до органел, можна вважати і складним молекулярним комплексом. Все частіше до органел зараховують і інші подібні комплекси порівняних розмірів та рівня складності - протеасоми,сплайсосоми та ін. У той же час порівняні за розмірами елементи цитоскелету (мікротрубочки, товсті філаменти поперечносмугастих м'язів і т. п.) зазвичай до органел не відносять.
Ступінь постійності клітинної структури - теж ненадійний критерій її віднесення до органел. Так, веретено поділу, що хоча і не постійно, але закономірно присутня у всіх еукаріотичних клітинах, зазвичай до органел не відносять, а везикули, які постійно з'являються і зникають в процесі обміну речовин - відносять. Багато в чому набір органоїдів, що перераховується до навчальних посібниках, визначається традицією.
Рибосома - найважливіший не мембранні органоїд живої клітини сферичної або злегка еліпсоїдної форми, діаметром 100-200 ангстрем, що складається з великої і малої субодиниць.
Рибосомами називають сферичні чи грибоподібні, дуже дрібні органели діаметром близько 22 нм. Вони були вперше описані і 1953 році американським біологом Д.Е.Паладе, який довів, що рибосоми являють собою нуклеопротеїди.
Кожна рибосома складається з двох субодиниць – великої і малої. Для характеристики рибосом використовують їхню властивість, як і всяких часток і молекул, осаджуватися з постійною швидкістю під дією відцентрової сили при диференційованому центрифугуванні. Досліди по седиментації виявили існування двох головних типів рибосом, що були названі 70S і 80S – рибосомами.
Перший тип являється у прокаріот, а більші 80 S –рибосомами – в еукаріотичних клітинах
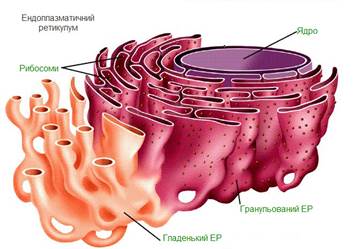
Рибосоми служать для біосинтезу білка з амінокислот по заданій матриці на основі генетичної інформації, що надається матричної РНК, або мРНК. Цей процес називається трансляцією. У еукаріотичних клітинах рибосоми розташовуються на мембранах ендоплазматичного ретикулума, хоча можуть бути локалізовані і в неприкріпленої формі в цитоплазмі. Нерідко з одного молекулою мРНК асоційоване кілька рибосом, така структура називається полірибосомами.
Синтез рибосом у еукаріот відбувається в спеціальній внутрішньоядерної структурі - ядерці. Рибосоми являють собою нуклеопротеїд, у складі якого відношення РНК/білок становить 1:1 у вищих тварин і 60-65:35-40 у бактерій.
До їх складу входять велика і мала субодиниці рибосомної РНК та білок.
Для стабілізації структури рибосом необхідні іони магнію. Кожна із субоди-ниць побудована з нуклеопротеїдного тяжа, де РНК взаємодіє з різними білка-ми і утворює тіло рибосоми, у яких амінокислоти сполучаються, тобто синтезу-ються білкові молекули. Кілька рибосом, зєднаних спільною ниткою інформа-ційної РНК, називають полісомами. Останні полісоми повязані з мембранами ендоплазматичної сітки, синтезують білки для виведення за межі клітини. Ступінь інтенсивності синтетичної діяльності вільних рибосом менший, а утворені білки використовуються для внутрішніх потреб клітини.
Мікрофіламенти. Це тонкі волоконця діаметром до 5 нм, побудовані з різних білків -- актину, міозину та ін. Вони розміщені переважно в кортикальній зоні клітини у складі її цитоплазматичних виростів і виконують роль цитоскелету, а також скоротливого апарату клітини.
Повні рибосомні частинки і їх субчастіції прийнято позначати відповідно до їх коефіцієнтами седиментації (швидкостями осадження, лат. Sedimentum - осад) в ультрацентрофугі, виражається в одиницях Сведберга (S). S - коефіцієнт седиментації, він залежить від молекулярної маси і просторової конформації частинки, осажджуючим при центрифугуванні.
Рибосомна РНК становить близько 70% всієї РНК клітини. Рибосоми еукаріот включають чотири молекули рРНК, з них 18S, 5.8S і 28S рРНК синтезуються в полісом РНК полімеразою I у вигляді єдиного попередника (45S), який потім піддається модифікаціям і нарізуванню. 5S рРНК синтезується РНК полімеразою III в іншій частині генома і не потребують додаткових модифікаціях.
Майже вся рРНК знаходиться у вигляді магнієвої солі, що необхідно для підтримки структури; при видаленні іонів магнію рибосома піддається дисоціації на субодиниці.
Рибосоми вперше були описані як ущільнені частки, або гранули, клітинним біологом румунського походження Джорджем Паладе в середині 1950-х років. У 1974 р. Паладе, Клод і Крістіан Де Дюв отримали Нобелівську премію з фізіології і медицини «за відкриття, що стосуються структурної і функціональної організації клітини».
Термін «рибосома» був запропонований Річардом Робертсом в 1958 замість «рібонуклеобелковая частинка мікросомальної фракції».
Біохімічні та мутаційні дослідження рибосоми починаючи з 1960-х дозволили описати багато функціональні та структурні особливості рибосоми.
В еукаріотичних організмах рибосоми можна знайти не лише в цитоплазмі, а й всередині в деяких великих мембранних органелах, зокрема в мітохондріях та хлоропластах.
Будова та молекулярний склад цих рибосом відрізняється від складу загально-клітинних рибосом, і є ближчим до складу рибосом прокаріотів. Такі рибосоми синтезують органело-специфічні білки, транслюючи органело-специфічну мРНК.
Вважається, що генетичні дефекти рибосомних білків та факторів біогенезу рибосом є летальними на ранніх ембріональних стадіях розвитку вищих організмів.
Експериментальний мутагенез рибосомних білків в Drosophila melanogaster (мутації minute) викликає спільний фенотип: занижена швидкість мітозу, зменшений розмір тіла, занижена фертильність, короткі війки.
Існує ряд свідчень що пов'язують раковутрансформацію клітин ссавців з розладами трансляційної системи загалом та розладами системи біогенезу рибосом зокрема.
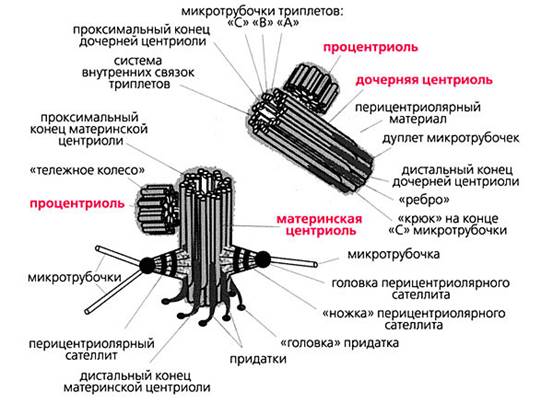
Центріоль - внутрішньоклітинний органоїд еукаріотічної клітини, що представляє тільця в структурі клітини, розмір яких знаходиться на кордоні роздільної здатності світлового мікроскопа. Ці органели в діляться клітинах беруть участь у формуванні веретена поділу і розташовуються на його полюсах.
У неподільних клітинах центріолі часто визначають полярність клітин епітелію і розташовуються поблизу комплексу Гольджі.
Термін був запропонований Теодором Бовері в 1895 році. Тонку будову центріолей вдалося вивчити за допомогою електронного мікроскопу.

Клітинний центр, або центросома, являє собою немембранний органоїд, локалізований біля ядра і складається з двох центріолей і центросферой. Причому постійним і найбільш важливим компонентом клітинного центру є центріолі. Цей органоид виявлений у клітинах тварин, нижчих рослин і грибів.
^ Центріолі (від лат. Centrum - серединна точка, центр) представляють два перпендикулярно розташованих один до одного циліндра, стінки яких утворені мікротрубочками і з'єднані системою зв'язувань.
Кінець одного циліндра (дочірня центриоль) спрямований до поверхні іншого (материнська центриоль). Сукупність зближених між собою материнської і дочірньої центріолей називаетя діплосомой.
Вперше центріолі були виявлені і описані в 1875 В. Флемінгом. У інтерфазних клітинах центріолі часто розташовуються біля комплексу Гольджі і ядра.
Ультрамікроскопічні будова центріолей було вивчено тільки за допомогою електронного мікроскопа. Стінку центріолей складають розташовані по колу 9 триплетів мікротрубочок, що утворюють порожнистий циліндр. Системи мікротрубочок центріолі можна описати формулою (9X3) + 0, підкреслюючи відсутність мікротрубочок в центральній частині.
Ширина центріолі становить близько 0,2 мкм, довжина - 0,3-0,5 мкм (проте, є центріолі, що досягають у довжину декількох мікрометрів).
Крім мікротрубочок до складу центріолі входять додаткові структури - "ручки", що з'єднують триплети.
Центросферой - щільний шар цитоплазми навколо центріолей, в якому часто містяться мікротрубочки, розташовані променями.
Центріолярний цикл.
Будова та активність центріолей змінюються залежно від періоду клітинного циклу. Це дозволяє говорити про центріолярном циклі.
На початку періоду G 1 від поверхні материнської центріолі починається ріст мікротрубочок, які ростуть і заповнюють цитоплазму.
У міру зростання мікротрубочки втрачають зв'язок з областю центріолей і можуть знаходитися в цитоплазмі тривалий час. У періоді S або G 2 відбувається подвоєння числа центріолей.
Цей процес полягає в тому, що центріолі в діплосоме розходяться і біля кожної з них відбувається закладка процентріолей. На початку поблизу і перпендикулярно вихідної центріолі закладаються дев'ять одиночних мікротрубочок.
Потім вони перетворюються в дев'ять дуплетів, а потім у дев'ять триплетів мікротрубочок нових центріолей. Цей спосіб збільшення числа центріолей був названий дуплікацією. Слід зазначити, що подвоєння числа центріолей не пов'язане з їх поділом, брунькуванням або фрагментацією, а відбувається шляхом утворення процентріолей.
Таким чином, в результаті дуплікації в клітці містяться чотири попарно зв'язані центріолі. У цьому періоді материнська центріоль продовжує відігравати роль центру освіти цитоплазматичних мікротрубочок. У періоді G 2 обидві материнські центріолі покриваються фібрилярні гало (зона тонких фібрил), від якого в профазі почнуть відростати мітотичні мікротрубочки. У цьому періоді в цитоплазмі відбувається зникнення мікротрубочок і клітка прагне придбати кулясту форму.
У профазі мітозу діплосоми розходяться до протилежних полюсі в клітини.
Від фібрилярного гало материнської центріолі відходять мікротрубочки, з яких формується веретено поділу мітотичного апарату.
Таким чином, центріолі є центрами організації росту мікротрубочок.
У телофазі відбувається руйнування веретена поділу. Слід зазначити, що в клітинах вищих рослин, деяких водоростей, грибів, ряду найпростіших центри організації росту мікротрубочок центріолей не мають.
У деяких найпростіших центрами індукції утворення мікротрубочок виступають щільні пластинки, пов'язані з мембраною.
У деяких об'єктах вдавалося спостерігати центріолі, зазвичай розташовані в парі (діплосома), і оточені зоною більш світлої цитоплазми, від якої радіально відходять тонкі фібрили (центросфери).
Сукупність центріолей і центросфери називають клітинним центром. Найчастіше пара центріолей лежить поблизу ядра.
Кожна центріоль побудована з циліндричних елементів (мікротрубочок), утворених в результаті полімеризації білкатубуліна. Дев'ять триплетів мікротрубочок розташовані по колу.
Центріолі беруть участь у формуванні цитоплазматичних мікротрубочок під час поділу клітини і в регуляції утворення мітотичного веретена. У клітинах вищих рослин і більшості грибів центріолей немає, і мітотичний веретено утворюється там іншим способом. Крім того, вчені вважають, що ферменти клітинного центру беруть участь у процесі переміщення дочірніх хромосом до різних полюсів в анафазі мітозу.
Мікротрубочки - це структури, в яких 13 тубулінових гетеродимерів укладені по окружності полого циліндра.
Мікротрубочки різного походження (війки найпростіших, клітини нервової тканини, веретено поділу) мають подібний склад і містять білки – тубуліну
. Очищені тубуліну при певних умовах здатні збиратися в мікротрубочки. Додавання алкалоїду колхіцину запобігає самосборку мікротрубочок або призводить до розбирання вже існуючих.
У клітці тубуліну існують у двох формах - вільної та зв'язаної. Зрушення рівноваги між цими формами призводить або до дисоціації мікротрубочок, або до їх зростання. Ні тубуліну в чистому вигляді, ні побудовані з них мікротрубочки не здатні до скорочення.
У клітинах мікротрубочки беруть участь у створенні низки тимчасових (цитоскелет інтерфазних клітин, веретено поділу) або постійних структур (центріолі, базальні тільця, війки і джгутики). Крім того, мікротрубочки сприяють орієнтованому руху внутрішньоклітинних компонентів.
Зовнішній діаметр циліндра близько 25 нм, внутрішній - близько 15. Один з кінців мікротрубочки, званий плюс-кінцем, постійно приєднує до себе вільний тубулін. Від протилежного кінця - мінус-кінця - тубулінових одиниці відщеплюються. В освіті мікротрубочки виділяють три фази: уповільнена фаза, або енуклеація.
Це етап зародження мікротрубочки, коли молекули тубулін починають з'єднуватися в більші утворення. Таке з'єднання відбувається повільніше, ніж приєднання тубулін до вже зібраної мікротрубочка, тому фаза і називається сповільненою; фаза полімеризації, або елонгація.
Мікротрубочки - білкові внутрішньоклітинні структури, що входять до складу цитоскелету. Мікротрубочки представляють собою порожні всередині циліндри діаметром 25 нм.
Довжина їх може бути від декількох мікрометрів до, ймовірно, декі.. лькох міліметрів в аксонах нервових клітин.
Їх стінка утворена димерами тубуліну. Мікротрубочки, подібно Актинові мікрофіламентам, полярні: на одному кінці відбувається самозбирання мікротрубочки, на іншому - розбирання.
У клітинах мікротрубочки грають роль структурних компонентів і беруть участь у багатьох клітинних процесах, включаючи мітоз, цитокінез і везикулярний транспорт.
Якщо концентрація вільного тубуліну висока, його полімеризація відбувається швидше, ніж деполімеризація на мінус-кінці, за рахунок чого мікротрубочка подовжується. У міру її росту концентрація тубуліну падає до критичної і швидкість росту сповільнюється аж до вступу в наступну фазу; фаза стабільного стану.
Деполімеризація врівноважує полімеризацію, і зростання мікротрубочки зупиняється. Лабораторні дослідження показують, що збірка мікротрубочок з тубулінів відбувається тільки в присутності гуанозінтріфосфата і іоновмагнія.
Мікротрубочки є динамічними структурами і в клітині постійно полімеризуються і деполімеризуються. Центросома, локалізована поблизу ядра, виступає в клітинах тварин і багатьох протистів як центр організації мікротрубочок (ЦОМТ): вони ростуть від неї до периферії клітини. Водночас мікротрубочки можуть раптово припинити своє зростання і вкоротитися назад у напрямку до Центросоми аж до повного руйнування, а потім вирости знову. При приєднанні до микротрубочка молекули тубуліну, що несуть ГТФ, утворюють «шапочку», яка забезпечує зростання мікротрубочки.
Якщо локальна концентрація тубуліну падає, пов'язана з бета-тубуліном ГТФ поступово гідролізується. Якщо повністю гідролізується ГТФ «шапочки» на ± кінці, це призводить до швидкого розпаду мікротрубочки. Таким чином, збирання й розбирання мікротрубочок пов'язана з витратами енергії ГТФ. Динамічна нестабільність мікротрубочок відіграє важливу фізіологічну роль.
Наприклад, при поділі клітини мікротрубочки ростуть дуже швидко і сприяють правильній орієнтації хромосом і утворення мітотичного веретена. Мікротрубочки в клітці використовуються як «рейок» для транспортування частинок. За їх поверхні можуть переміщатися мембранні бульбашки і мітохондрії. Транспортування по микротрубочкам здійснюють білки, звані моторними. Це високомолекулярні сполуки, які з двох важких (масою близько 300 кДа) і декількох легких ланцюгів.
У важких ланцюгах виділяють головний і хвостовій домени. Два головних домену зв'язуються з мікротрубочками і є власне двигунами, а хвостові - зв'язуються з органелами та іншими внутрішньоклітинними утвореннями, такими, що підлягають транспортуванню. Виділяють два види моторних білків: цитоплазматичні динеін; кінезина.
Головні домени моторних білків для цього містять АТФ-звязуючі ділянки. Типічна аксонема містить 9 пароб*єднаних мікротрубочок по периферії і дві повні мікротрубочки в центрі
Із мікротрубочок складаються також центріолі та веретеноподіли, що забезпечують розходження хромосом до полюсів клітин при мітозі і мейозі. Мікротрубочки беруть участь в підтриманні форми клітини в роміщенні органоїдів.
Їх властивості динаміки сильно відрізняються від відповідних характеристик мікротрубочок, такіактинових філаментів.
Список використаної літератури, посилання
1.Яригин Н.В., Васильєва В.І., Волков І. М., Синельщиков В.В. Біологія.Учеб. для мед. спец. вузів.Т-1,Т-2. - Москва, 2004. – 431 з.
2. Крістіан ДеДюв. Подорож у світ живою клітиною. -256 з.
3. Медична біологія: посібник з практичних занять О.В. Романенко, М. І. Кравчук, В. І. Гринченко та ін.; За редакцією О. В. Романенка.- К.:Здоров’я. 2005, с. 305
4.Слюсарєв А.О., Жукова С.В. Біологія: Біологія: Підручник / Пер. З рос. В. О. Мотузний.- К.: Вища школа,1992. С. – 422
5. Медична біологія: Підручник / за ред. В. П. Пішака, Ю.В. Бажори. – Видання 2-ге перероблене і доповнене. – Вінниця: НОВА КНИГА А. 2009 с. - 608

