 2018-01-21
2018-01-21 1576
1576Рассматривая макроструктуру скелетных мышц, были выделены три основных элемента: это фасции, мышечные волокна и сухожилия. В этом параграфе мы более подробно рассмотрим типы мышечных волокон, так как от этого во многом зависит способность мышц к проявлению силы, скорости, а также выносливости.
Вначале были выделены два типа скелетных мышечных волокон, каждый из которых имел свои физиологические особенности. Это – медленные (тонические) и быстрые (фазические) волокна. В некоторых мышцах могут быть только быстрые или только медленные волокна, в других – волокна обоих типов в определенном соотношении. В дальнейшем были выделены мышечные волокна промежуточного типа.
Благодаря различным типам волокон организм способен передвигаться и поддерживать позу. Быстрые волокна позволяют мышце сокращаться с высокой скоростью. В большом количестве эти волокна имеются у хищников; они обеспечивают быстроту реакции при ловле добычи. С другой стороны, потенциальная добыча, чтобы не стать жертвой хищников, тоже должна быть способна к быстрому реагированию. В обоих случаях от подвижности животного будут зависеть его шансы на выживание. Когда животное находится в покое, оно поддерживает определенную позу с помощью тонических мышечных волокон. Им свойственно более медленное и длительное сокращение и в то же время энергетические затраты меньше, чем при сокращении быстрых волокон. Сокращение их по своей природе обычно изометрическое, при котором мышцы, противодействуя силе тяжести и удерживая конечности в определенном положении, сохраняют постоянную длину.
У человека все мышцы тела состоят из волокон трех типов, но обычно один из них доминирует. Это имеет физиологическое значение, поскольку тонические мышцы способны к медленному и длительному сокращению и поэтому медленных волокон больше в позных мышцах-разгибателях, тогда как в сгибателях, предназначенных для быстрых реакций, преобладают быстрые фазические волокна.
Согласно современным представлениям, большинство скелетных мышц человека и животных представляют собой гетерогенные морфофункциональные системы, состоящие из мышечных волокон, отличающихся по структуре, метаболизму и функции.
Мышечные волокнаIтипа в мировой номенклатуре обозначают как красные, окислительные, медленные, устойчивые к утомлению. В мышечных волокнах I типа хорошо выражен Z-диск, который толще, чем в мышечных волокнах других типов, саркоплазмаческийретикулум развит в меньшей степени, чем в других типах мышечных волокон. В данных структурах выявляется много митохондрий с поперечной ориентацией. В саркоплазме обнаруживаются липидные капли, которые часто контактируют с митохондриями. Мышечные волокна I типа характеризуются также высокой степенью кровообращения. Каждое мышечное волокно I типа контактирует с 5-6 кровеносными капиллярами. В этих волокнах отмечается самое высокое содержание миоглобина. Согласно данным гистохимических исследований, в волокнах I типа обнаруживается более низкая активность креатинфосфокиназы, чем в других мышечных волокнах.
Мышечные волокнаIIВ типа в мировой номенклатуре обозначают как белые, быстрые, гликолитические, быстроутомляемые. Z-диск в мышечных волокнах IIВ типа более тонкий, чем в волокнах других типов. В данных структурах очень хорошо развит саркоплазматический ретикулум, системы терминальных цистерн и триад. В саркоплазме мышечных волокон IIB типа сравнительно немного малых по размерам митохондрий, содержащих небольшое число крист. Мышечные волокна IIВ типа контактируют с меньшим количеством капилляров, чем мышечные волокна других типов. Кроме того, в этих волокнах имеется очень мало миоглобина. Именно этим объясняется их более светлая окраска, благодаря чему они и получили название «белых». В мышечных волокнах IIB типа содержание креатинфосфата выше, чем в других типах мышечных волокон.
Мышечные волокнаIIA типа в мировой номенклатуре обозначают как промежуточные, окислительно-гликолитические, быстрые, устойчивые к утомлению. Z-диск в мышечных волокна IIA типа, по мнению ряда авторов, толще, чем в мышечных волокнах IIВ типа и не отличается от такового в волокнах I типа. По данному показателю волокна типа IIA занимают промежуточное положение между мышечными волокнами I типа и мышечными волокнами IIB типа. В мышечных волокнах IIA типа хорошо развит саркоплазматический ретикулум и системы триад. Мышечные волокна IIА типа богаче митохондриями, чем волокна IIВ типа и практически не отличаются по этому признаку от мышечных волокон I типа. Митохондрии в мышечных волокнах I типа имеют преимущественно продольную ориентацию (по длине оси мышечных волокон). Они часто образуют скопления под сарколеммой. В саркоплазме мышечных волокон IIA типа встречается достаточно большое количество липидных капель. Мышечные волокна IIА типа содержат больше миоглобина, чем мышечные структуры IIВ типа. Содержание креатинфосфата в мышечных волокнах IIА типа ниже, чем в волокнах IIВ типа и не отличается от такового в волокнах I типа. Мышечные волокна данного типа представляются универсальными в плане их метаболизма. В связи с тем, что мышечные волокна IIА типа способны длительное время производить работу достаточно большой мощности, они должны рассматриваться как универсальные мышечные волокна в плане их функциональных возможностей.
8. Механизм образования мочи: клубочковая фильтрация, канальцеваяреабсорбция и секреция. Значение петли Генле в образовании конечной мочи.
Клубочковая фильтрация. Начальным этапом образвания мочи является фильтрация: в почечном тельце из капиллярного клубочка в полость капсулы фильтруется жидкая часть крови. Клубочковая фильтрация – это пассивный процесс. В условиях покоя у взрослого человека около 1/4 крови, выбрасываемой в аорту левым желудочком сердца, поступает в почечные артерии. Иными словами, через обе почки у взрослого мужчины проходит около 1300 мл крови в минуту, у женщин несколько меньше. Общая фильтрационная поверхность клубочков почек составляет примерно 1,5 м2. В клубочках из кровеносных капилляров в просвет капсулы почечного клубочка происходит ультрафильтрация плазмы крови, в результате чего образуется первичная моча, в которой практически отсутствует белок. В норме белки как коллоидныевещества не проходят через стенку капилляров в полость капсулы почечного клубочка. При ряде патологических состояний проницаемость мембраны почечного фильтра повышается, что ведет к изменению состава ультрафильтрата. Повышение проницаемости является главной причиной протеинурии, прежде всего альбуминурии. В норме объемная скорость фильтрации в среднем составляет 125 мл/мин, что в 100 раз превышает продукцию конечной мочи. Скорость фильтрации обеспечивается фильтрационным давлением, которое можно выразить следующей формулой:
ФД = КД – (ОД + КапсД),
где ФД – фильтрационное давление; КД – капиллярное давление; ОД – онко-тическое давление; КапсД – внутрикапсулярное давление. Следовательно, для обеспечения процесса фильтрации необходимо, чтобы гидростатическое давление крови в капиллярах превышало сумму онкотического и внутрикап-сулярного. В норме эта величина составляет около 40 гПа (30 мм рт. ст.). Вещества, усиливающие кровообращение в почках или увеличивающие количество функционирующих клубочков (например, теобромин, теофил-лин, плоды можжевельника, листья толокнянки и др.), обладают мочегонными свойствами.
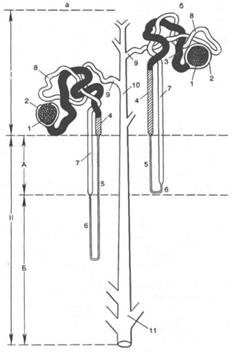
Рис. 18.1. Строение юкстамедуллярного (а) и коркового (б) нефронов. I - корковое вещество; II - мозговое вещество; А - наружная зона мозгового вещества; Б - внутренняя зона мозгового вещества; 1 - сосудистый клубочек; 2 - капсула почечного клубочка; 3 - проксимальный каналец (извитая часть); 4 - проксимальный каналец (прямая часть); 5 - нисходящее тонкое колено петли нефрона; 6 - восходящее тонкое колено петли нефрона; 7 - восходящее толстое колено петли нефрона; 8 - дистальный извитой каналец; 9 - связующий каналец; 10 - собирательная трубка; 11 - собирательная почечная трубочка.
Капиллярное давление в почках зависит не столько от артериального давления, сколько от соотношения просвета «приносящей» и «выносящей» артериол клубочка. «Выносящая» артериола примерно на 30% меньше в диаметре, чем «приносящая», регуляция их просвета осуществляется прежде всего кининовой системой. Сужение «выносящей» артериолы увеличивает фильтрацию. Напротив, сужение «приносящей» артериолы снижает фильтрацию.
По величине клубочковой фильтрации судят о фильтрационной способности почек. Если в кровяное русло ввестивещество, которое фильтруется в клубочках, но не реабсорбируется и не секретируется канальцами нефро-нов, то его клиренс численно равен объемной скорости клубочковой фильтрации. Клиренс (очищение) любого соединения принято выражать количеством миллилитров плазмы, которое в 1 мин полностью освобождается от определенноговещества при прохождении ее через почки. Веществами, по которым чаще определяют клубочковую фильтрацию, являются инулин и маннитол. Для расчета клиренса (например, инулина) необходимо величину минутного диуреза умножить на Kм/Kкp(отношение концентраций данного вещества в моче и плазме крови):

где С – клиренс; Км – концентрация данного соединения в моче; Ккр – концентрация в плазме крови; V – количествомочи в 1 мин, мл. Например, при расчете клиренса инулина в норме получим величину клубочковой фильтрации, равную 100–125 мл за 1 мин.
Реабсорбция и секреция. Суточное количество ультрафильтрата в 3 раза превышает общее количество жидкости, содержащейся в организме. Естест-
венно, что первичная моча во время движения по почечным канальцам отдает большую часть своих составных частей, особенно воду, обратно в кровь. Лишь 1 % жидкости, профильтрованной клубочками, превращается в мочу.
В канальцах реабсорбируется 99% воды, натрия, хлора, гидрокарбоната, аминокислот, 93% калия, 45% мочевины и т.д. Из первичной мочи в результате реабсорбции образуется вторичная, или окончательная, моча, которая затем поступает в почечные чашки, лоханку и по мочеточникам попадает в мочевой пузырь.
Функциональное значение отдельных почечных канальцев в процессе мочеобразования неодинаково. Клеткипроксимального сегмента нефрона реабсорбируют попавшие в фильтрат глюкозу, аминокислоты, витамины,электролиты; 6/7 жидкости, составляющей первичную мочу, подвергается реабсорбции также в проксимальных канальцах. Вода первичной мочи частично (парциально) реабсорбируется в дистальных канальцах. В этих же канальцах происходит дополнительнаяреабсорбция натрия, могут секрети-роваться в просвет нефрона ионы калия,аммония, водорода и др.
В настоящее время в значительной степени изучены молекулярные механизмы реабсорбции и секреции веществклетками почечных канальцев. Так, установлено, что при реабсорбции натрий пассивно поступает из просвета канальца внутрь клетки, движется по ней к области базальной плазматической мембраны и с помощью «натриевогонасоса» поступает во внеклеточную жидкость. До 80% энергии АТФ в клетках канальцев почек расходуется на «натриевый насос». Всасывание воды в проксимальном сегменте происходит пассивно в результате активного всасывания натрия. Вода в этом случае «следует» за натрием. Кстати, в дистальном сегменте всасывание водыпроисходит вне всякой зависимости от всасывания ионов натрия; этот процесс регулируется антидиуретическим гормоном.
Калий в отличие от натрия может не только реабсорбироваться, но и секретироваться. При секреции калий из межклеточной жидкости поступает через базальную плазматическую мембрану в клетку канальца за счет работы «натрий-калиевого насоса», а затем выделяется в просвет нефрона через апикальную клеточную мембранупассивно. Секреция, как и реабсорб-ция, является активным процессом, связанным с функцией клеток канальцев. Механизмы секреции те же, что и механизмы реабсорбции, но только все процессы протекают в обратном направлении – от крови к канальцу.
Вещества, которые не только фильтруются через клубочки, но и реабсор-бируются или секретируются в канальцах, имеют клиренс, который показывает целостную работу почек (смешанный клиренс). В зависимости от того, комбинируется ли фильтрация с реабсорбцией или с секрецией, выделяют два вида смешанного клиренса: фильтрационно-реабсорбционный и фильт-рационно-секреционный. Величина смешанного фильтрационно-реабсорб-ционного клиренса меньше величины клубочкового клиренса, так как часть вещества реабсорбируется из первичной мочи в канальцах. Значение этого показателя тем меньше, чем эффективнее реабсорбция в канальцах. Так, для глюкозы в норме он равен 0. Максимальное всасывание глюкозы в канальцах составляет 350 мг/мин. Максимальную способность канальцев к обратному всасыванию принято обозначать Тм (транспорт максимум). Иногда встречаются пациенты с заболеванием почек, которые, несмотря на высокое содержание глюкозы в плазме крови, не выделяют глюкозу с мочой, так как фильтруемое количество глюкозы ниже значения Тм. Наоборот, при врожденном заболевании почечнаяглюкозурия может быть основана на снижении значения Тм.
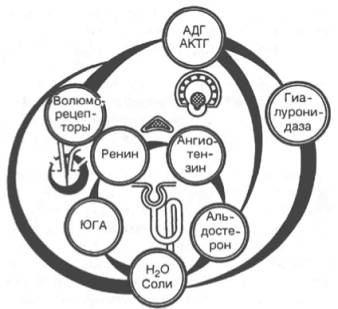
Рис. 18.2. Регуляция реабсорбции в почке (схема по А.П. Зильберу). Объяснение в тексте.
Для мочевины величина смешанного фильтрационно-реабсорбционного клиренса составляет 70. Это значит, что из каждых 125 мл ультрафильтрата или плазмы крови за минуту от мочевины полностью освобождаются 70 мл. Иными словами, определенное количество мочевины, а именно то, которое содержится в 55 мл ультрафильтратаилиплазмы, всасывается обратно.
Величина смешанного фильтрационно-секреционного клиренса может быть больше клубочкового клиренса, так как к первичной моче прибавляется дополнительное количество вещества, которое секретируется в канальцах. Этот клиренс тем больше, чем сильнее секреция канальцев. Клиренс некоторых веществ, секретируемых канальцами (например, диодраст, пара-аминогиппуровая кислота), настолько высок, что практически приближается к величине почечного кровотока (количество крови, которое за минуту проходит через почки). Таким образом, по клиренсу этихвеществ можно определить величину кровотока.
Реабсорбция и секреция различных веществ регулируются ЦНС и гормональными факторами. Например, при сильных болевых раздражениях или отрицательных эмоциях может возникнуть анурия (прекращение процесса мочеобразования). Всасывание воды возрастает под влиянием антидиуретического гормона вазопрессина. Альдостерон увеличивает реабсорбцию натрия в канальцах, а вместе с ним и воды. Всасывание кальция и фосфатаизменяется под влиянием паратиреоидного гормона. Паратгормон стимулирует секрецию фосфата, а витамин D задерживает ее.
Регуляция реабсорбции натрия и воды в почке представлена на рис. 18.2. При недостаточном поступлении крови к почечным клубочкам, сопровождающемся небольшим растяжением стенок артериол (снижение давления), происходит возбуждение заложенных в стенках артериол клеток юкстагло-мерулярного аппарата (ЮГА). Они начинают усиленно секретировать протеолитический фермент ренин, катализирующий начальный этап образования ангиотензина. Субстратом ферментативного действия ренина является ангиотензиноген (гликопротеин), относящийся к α2-глобулинам и содержащийся в плазме крови и лимфе.
Ренин разрывает в молекуле ангиотензиногена пептидную связь, образованную двумя остатками лейцина, в результате чего освобождается дека-пептид ангиотензин I, биологическая активность которого незначительна в среде, близкой к нейтральной.
Считают, что под влиянием специальной пептидазы, обнаруженной в плазме крови и тканях,– ангиотензин I превращающего фермента (дипеп-тидил-карбоксипептидаза I) из ангиотензина I образуется октапептид ан-гиотензин II. Главным местом этого превращения являются легкие.
Петля Генле состоит из нисходящей и восходящей частей. Тонкий сегмент нисходящей части является продолжением проксимального канальца и опускается из коркового вещества почки в мозговое. В мозговом веществе тонкий сегмент загибается, делая U-образный разворот, и поднимается в корковое вещество уже как восходящая часть петли Генле. В восходящей части выделяют различающиеся в функциональном отношении тонкий сегмент, толстый медуллярный сегмент и толстый кортикальный сегмент (рис. 31-1). У нефронов, клубочки которых расположенные вблизи мозго-
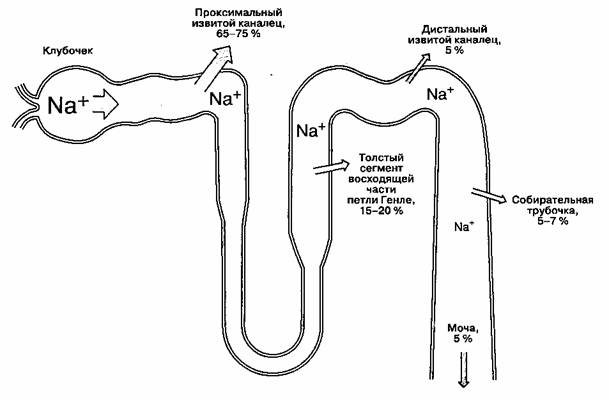
Рис. 31-2. Реабсорбция натрия в нефроне. Указанное количество реабсорбированного натрия в каждом отделе нефро-на соответствует проценту от профильтровавшейся натриевой нагрузки. (Из: Cogan M. G. Fluid and Electrolytes: Physiology and Pathophysiology, 14th ed. Appleton&Lange, 1991; с разрешения.)
вого вещества (юкстамедуллярные нефроны), петля Генле более длинная, чем у нефронов, клубочки которых лежат ближе к наружной поверхности почки (корковые нефроны). У корковых нефронов с короткой петлей отсутствует тонкий сегмент восходящей части. Корковых нефронов в 7 раз больше, чем юкстамедуллярных. Петля Генле поддерживает гипертоничность интерстициальной жидкости мозгового вещества, а также опосредованно обеспечивает процесс концентрирования мочи в собирательных трубочках.
В норме до 25-35 % фильтрата, образующегося в капсуле Боумена, достигает петли Генле. В ней реабсорбируется 15-20% фильтруемого натрия. За исключением толстого сегмента восходящей части, реабсорбция растворенных веществ и воды в петле Генле происходит пассивно — по градиенту концентрации и осмотическому градиенту соответственно. В толстом сегменте восходящей части Na" и СГ реабсорбируются в большей степени, чем вода; более того, в этой части нефрона реабсорбцияNa" непосредственно сопряжена с реабсорбцией K+ и СГ (рис. 31-4), и концентрация СГ в канальцевой жидкости является фактором, ограничивающим
скорость реабсорбции. Активная реабсорбцияNa+ осуществляется Ка"УК+-зависимой АТФ-азой капиллярной поверхности эпителиальных клеток.
В отличие от нисходящей части и тонкого сегмента восходящей части петли Генле, толстый сегмент восходящей части непроницаем для воды. Поэтому оттекающая из петли Генлеканальцевая жидкость гипотонична (100-200 мОсм/л), а окружающая петлю Генле интерстициальная жидкость гипертонична. Механизм противоточного умножения работает таким образом, что гипертонич-ностъканалъцевой жидкости и окружающего UH-терстиция значительно нарастает по мере углубления в мозговое вещество почки (рис. 31-5). Концентрация мочевины в мозговом веществе становится высокой, что существенно влияет наегогипертоничность. Структуры механизма противоточного умножения включают петлю Генле, кортикальные и медуллярные собирательные трубочки и сопровождающие их капилляры (vasarecta),
Толстый сегмент восходящей части петли Генле играет важную роль в реабсорбции Ca2+ и Mg2+.
9. Механизм мышечного сокращения. Роль саркоплазматического ретикулума в процессах сокращения и расслабления мышечного волокна.
Мы́шечноесокраще́ние — реакция мышечных клеток на воздействие нейромедиатора, реже гормона, проявляющаяся в уменьшении длины клетки. Это жизненно важная функция организма, связанная с оборонительными, дыхательными, пищевыми, половыми, выделительными и другими физиологическими процессами.
Все виды произвольных движений — ходьба, мимика, движения глазных яблок, глотание, дыхание и т. п. осуществляются за счёт скелетных мышц. Непроизвольные движения (кроме сокращения сердца) — перистальтика желудка и кишечника, изменение тонуса кровеносных сосудов, поддержание тонуса мочевого пузыря — обусловлены сокращением гладкой мускулатуры. Работа сердца обеспечивается сокращением сердечной мускулатуры.
Основой всех типов мышечного сокращения служит взаимодействие актина и миозина. В скелетных мышцах за сокращение отвечаютмиофибриллы (примерно две трети сухого веса мышц). Миофибриллы — структуры толщиной 1 — 2 мкм, состоящие из саркомеров — структур длиной около 2,5 мкм, состоящих из актиновых и миозиновых (тонких и толстых) филаментов и Z-дисков, соединённых с актиновымифиламентами. Сокращение происходит при увеличении концентрации в цитоплазме ионов Ca2+ в результате скольжения миозиновыхфиламентов относительно актиновых. Источником энергии сокращения служит АТФ. КПД мышечной клетки около 50 %, мышцы в целом не более 20%. Максимальная сила мышц не достигается в реальных условиях; не все клетки мышцы используются одновременно и сокращаются с максимальной силой, иначе при сокращении многих скелетных мышц будут повреждены сухожилия или кости (что иногда и наблюдается при сильных судорогах). КПД мышцы также зависит от внешних условий; например, на холоде он значительно снижается, так как для организма важнее сохранить температуру тела.[1]
Скольжение миозина относительно актина[править | править вики-текст]
Головки миозина расщепляют АТФ и за счет высвобождающейся энергии меняют конформацию, скользя по актиновымфиламентам. Цикл можно разделить на 4 стадии:
1. Свободная головка миозина связывается с АТФ и гидролизует его до АДФ и фосфата и остаётся связанной с ними. (Обратимый процесс — энергия, выделившаяся в результате гидролиза, запасается в изменённой конформации миозина).
2. Головки слабо связываются со следующей субъединицей актина, фосфат отделяется, и это приводит к прочному связыванию головки миозина с актиновымфиламентом. Эта реакция уже необратима.
3. Головка претерпевает конформационное изменение, производящее подтягивание толстогофиламента к Z-диску (или, что эквивалентно, свободных концов тонких филаментов друг к другу).
4. Отделяется АДФ, за счёт этого головка отделяется от актиновогофиламента. Присоединяется новая молекула АТФ.
Далее цикл повторяется до уменьшения концентрации ионов Ca2+ или исчерпании запаса АТФ (в результате смерти клетки). Скорость скольжения миозина по актину ≈15 мкм/сек. В миозиновомфиламенте много (около 500) молекул миозина и, следовательно, при сокращении цикл повторяется сотнями головок сразу, что и приводит к быстрому и сильному сокращению. Следует заметить, что миозиин ведёт себя как фермент — актин-зависимаяАТФаза. Так как каждое повторение цикла связано с гидролизом АТФ, а следовательно, с положительным изменением свободной энергии, то процесс однонаправленный. Миозин движется по актину только в сторону плюс-конца.
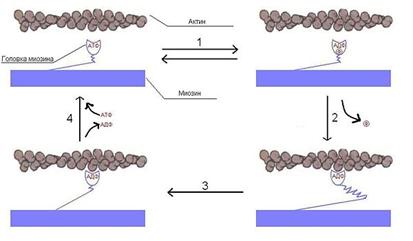
Последовательные стадии
Источник энергии для сокращения[править | править вики-текст]
Для сокращения мышцы используется энергия гидролиза АТФ, но мышечная клетка имеет крайне эффективную систему регенерации запаса АТФ, так что в расслабленной и работающей мышце содержание АТФ примерно равно. Фермент фосфокреатинкиназа катализирует реакцию между АДФ и креатинфосфатом, продукты которой — АТФ и креатин. Креатинфосфат содержит больше запасённой энергии, чем АТФ. Благодаря этому механизму при вспышке активности в мышечной клетке падает содержание именно креатинфосфата, а количество универсального источника энергии — АТФ — не изменяется. Механизмы регенерации запаса АТФ могут различаться в зависимости от парциального давления кислорода в окружающих тканях (см. Анаэробные организмы).
Механизм регуляции[править | править вики-текст]
В основном в регуляции мышечной активности участвуют нейроны, но есть случаи, когда сокращением гладкой мускулатуры управляют и гормоны (например, адреналин и окситоцин). Сигнал о сокращении можно разделить на несколько этапов:
От клеточной мембраны до саркоплазматического ретикулума[править | править вики-текст]
Воздействие медиатора, выделившегося из мотонейрона, вызывает потенциал действия на клеточной мембране мышечной клетки, который передаётся далее с помощью специальныхвпячиваний мембраны, называемых Т-трубочками, которые отходят от мембраны внутрь клетки. От Т-трубочек сигнал передаётся саркоплазматическомуретикулуму — особому компартменту из уплощенных мембранных пузырьков (эндоплазматической сети мышечной клетки), окружающих каждую миофибриллу. Этот сигнал вызывает открытие Ca2+-каналов в мембране ретикулума. Обратно ионы Ca2+попадают в ретикулум с помощью мембранных кальциевых насосов — Ca2+-АТФазы.
От выделения ионов Ca2+ до сокращения миофибрилл[править | править вики-текст]
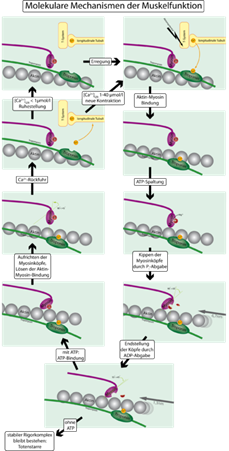
Механизм сокращения мышц с учётом тропонина и тропомиозина
Для того, чтобы контролировать сокращение, к актиновомуфиламенту прикрепляется белок тропомиозин и комплекс из трёх белков —тропонин (субъединицы этого комплекса называются тропонинами T,I и C). Тропонин C — близкий гомолог другого белка,кальмодулина. Через каждые семь субъединиц актина расположен только один тропониновый комплекс. Связь актина с тропонином I перемещает тропомиозин в положение, мешающее связи миозина с актином. Тропонин C связывается с четырьмя ионами Ca2+ и ослабляет действие тропонина I на актин, и тропомиозин занимает положение, не препятствующее связи актина с миозином. Источником энергии для сокращения мышечных волокон служит АТФ. При связывании тропонина с ионами кальция активируются каталитические центры для расщепления АТФ на головках миозина. За счет ферментативной активности головок миозина гидролизуется АТФ, расположенный на головке миозина, что обеспечивает энергией изменение конформации головок и скольжение нитей. Освобождающиеся при гидролизе АТФ молекула АДФ и неорганический фосфат используются для последующего ресинтеза АТФ. К миозиновой головке присоединяется новая молекула АТФ. При этом происходит разъединение поперечного мостика с нитью актина. Повторное прикрепление и отсоединение мостиков продолжается до тех пор, пока концентрация кальция внутри миофибрилл не снизится до подпороговой величины. Тогда мышечные волокна начинают расслабляться.
10. Нейрон как структурная и функциональная единица нервной системы. Классификация нейронов. Строение нейрона, функции отдельных частей нейрона. Нейроглия, её значение.
Нейрон (от др.-греч. νεῦρον — волокно, нерв) — это структурно-функциональная единица нервной системы. Эта клетка имеет сложное строение, высокоспециализирована и по структуре содержит ядро, тело клетки и отростки. В организме человека насчитывается более восьмидесяти пяти миллиардов нейронов.
Тело клетки[править | править вики-текст]
Тело нервной клетки состоит из протоплазмы (цитоплазмы и ядра), снаружи ограничена мембраной из двойного слоя липидов (билипидный слой). Липиды состоят из гидрофильных головок и гидрофобных хвостов, расположены гидрофобными хвостами друг к другу, образуя гидрофобный слой, который пропускает только жирорастворимые вещества (напр. кислород и углекислый газ). На мембране находятся белки: на поверхности (в форме глобул), на которых можно наблюдать наросты полисахаридов (гликокаликс), благодаря которым клетка воспринимает внешнее раздражение, и интегральные белки, пронизывающие мембрану насквозь, в которых находятся ионные каналы.
Нейрон состоит из тела диаметром от 3 до 130 мкм, содержащего ядро (с большим количеством ядерных пор) и органеллы (в том числе сильно развитый шероховатый ЭПР с активными рибосомами, аппарат Гольджи), а также из отростков. Выделяют два вида отростков: дендриты и аксон. Нейрон имеет развитый и сложный цитоскелет, проникающий в его отростки. Цитоскелет поддерживает форму клетки, его нити служат «рельсами» для транспорта органелл и упакованных в мембранные пузырьки веществ (например, нейромедиаторов). Цитоскелет нейрона состоит из фибрилл разного диаметра: Микротрубочки (Д = 20-30 нм) — состоят из белка тубулина и тянутся от нейрона по аксону, вплоть до нервных окончаний. Нейрофиламенты (Д = 10 нм) — вместе с микротрубочками обеспечивают внутриклеточный транспорт веществ. Микрофиламенты (Д = 5 нм) — состоят из белков актина и миозина, особенно выражены в растущих нервных отростках и в нейроглии. В теле нейрона выявляется развитый синтетический аппарат, гранулярная ЭПС нейрона окрашивается базофильно и известна под названием «тигроид». Тигроид проникает в начальные отделы дендритов, но располагается на заметном расстоянии от начала аксона, что служит гистологическим признаком аксона. Нейроны различаются по форме, числу отростков и функциям. В зависимости от функции выделяют чувствительные, эффекторные(двигательные, секреторные) и вставочные. Чувствительные нейроны воспринимают раздражения, преобразуют их в нервные импульсы и передают в мозг. Эффекторные (от лат.эффектус — действие) — вырабатывают и посылают команды к рабочим органам. Вставочные — осуществляют связь между чувствительными и двигательными нейронами, участвуют в обработке информации и выработке команд.
Различается антероградный (от тела) и ретроградный (к телу) аксонный транспорт.
Дендриты и аксон[править | править вики-текст]
Основные статьи: Дендрит, Аксон
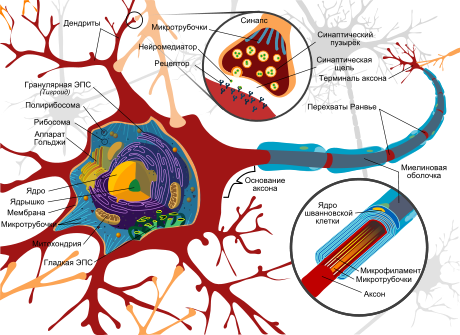
Схема строения нейрона
Аксон — обычно длинный отросток нейрона, приспособленный для проведения возбуждения и информации от тела нейрона или от нейрона к исполнительному органу. Дендриты — как правило, короткие и сильно разветвлённые отростки нейрона, служащие главным местом образования влияющих на нейрон возбуждающих и тормозных синапсов (разные нейроны имеют различное соотношение длины аксона и дендритов), и которые передают возбуждение к телу нейрона. Нейрон может иметь несколько дендритов и обычно только один аксон. Один нейрон может иметь связи со многими (до 20 тысяч) другими нейронами.
Дендриты делятся дихотомически, аксоны же дают коллатерали. В узлах ветвления обычно сосредоточены митохондрии.
Дендриты не имеют миелиновой оболочки, аксоны же могут её иметь. Местом генерации возбуждения у большинства нейронов является аксонный холмик — образование в месте отхождения аксона от тела. У всех нейронов эта зона называется триггерной.
Синапс[править | править вики-текст]
Основная статья: Синапс
Си́напс (греч. σύναψις, от συνάπτειν — обнимать, обхватывать, пожимать руку) — место контакта между двумя нейронамиили между нейроном и получающей сигнал эффекторной клеткой. Служит для передачи нервного импульса между двумя клетками, причём в ходе синаптической передачи амплитуда и частота сигнала могут регулироваться. Одни синапсывызывают деполяризацию нейрона, другие — гиперполяризацию; первые являются возбуждающими, вторые — тормозными. Обычно для возбуждения нейрона необходимо раздражение от нескольких возбуждающих синапсов.
Термин был введён в 1897 г. английским физиологом Чарльзом Шеррингтоном.
Классификация[править | править вики-текст]
Структурная классификация[править | править вики-текст]
На основании числа и расположения дендритов и аксона нейроны делятся на безаксонные, униполярные нейроны, псевдоуниполярные нейроны, биполярные нейроны и мультиполярные (много дендритных стволов, обычно эфферентные) нейроны.
Безаксонные нейроны — небольшие клетки, сгруппированы вблизи спинного мозга в межпозвоночных ганглиях, не имеющие анатомических признаков разделения отростков на дендриты и аксоны. Все отростки у клетки очень похожи. Функциональное назначение безаксонных нейронов слабо изучено.
Униполярные нейроны — нейроны с одним отростком, присутствуют, например в сенсорном ядре тройничного нерва в среднем мозге. Многие морфологи считают, что униполярные нейроны в теле человека и высших позвоночных не встречаются.
Биполярные нейроны — нейроны, имеющие один аксон и один дендрит, расположенные в специализированных сенсорных органах — сетчатке глаза, обонятельном эпителии и луковице, слуховом и вестибулярном ганглиях.
Мультиполярные нейроны — нейроны с одним аксоном и несколькими дендритами. Данный вид нервных клеток преобладает в центральной нервной системе.
Псевдоуниполярные нейроны — являются уникальными в своём роде. От тела отходит один отросток, который сразу же Т-образно делится. Весь этот единый тракт покрыт миелиновой оболочкой и структурно представляет собой аксон, хотя по одной из ветвей возбуждение идёт не от, а к телу нейрона. Структурно дендритами являются разветвления на конце этого (периферического) отростка. Триггерной зоной является начало этого разветвления (то есть находится вне тела клетки). Такие нейроны встречаются в спинальных ганглиях.
Функциональная классификация[править | править вики-текст]
По положению в рефлекторной дуге различают афферентные нейроны (чувствительные нейроны), эфферентные нейроны (часть из них называется двигательными нейронами, иногда это не очень точное название распространяется на всю группу эфферентов) и интернейроны (вставочные нейроны).
Афферентные нейроны (чувствительный, сенсорный, рецепторный или центростремительный). К нейронам данного типа относятся первичные клетки органов чувств и псевдоуниполярные клетки, у которых дендриты имеют свободные окончания.
Эфферентные нейроны (эффекторный, двигательный, моторный или центробежный). К нейронам данного типа относятся конечные нейроны — ультиматные и предпоследние — не ультиматные.
Ассоциативные нейроны (вставочные или интернейроны) — группа нейронов осуществляет связь между эфферентными и афферентными, их делят на интризитные, комиссуральные и проекционные.
Секреторные нейроны — нейроны, секретирующие высокоактивные вещества (нейрогормоны). У них хорошо развит комплекс Гольджи, аксон заканчивается аксовазальными синапсами.
Морфологическая классификация[править | править вики-текст]
Морфологическое строение нейронов многообразно. В связи с этим при классификации нейронов применяют несколько принципов:
· учитывают размеры и форму тела нейрона;
· количество и характер ветвления отростков;
· длину аксона и наличие специализированных оболочек.
По форме клетки, нейроны могут быть сферическими, зернистыми, звездчатыми, пирамидными, грушевидными, веретеновидными, неправильными и т. д. Размер тела нейрона варьирует от 5 мкм у малых зернистых клеток до 120—150 мкм у гигантских пирамидных нейронов.
По количеству отростков выделяют следующие морфологические типы нейронов[1]:
· униполярные (с одним отростком) нейроциты, присутствующие, например, в сенсорном ядре тройничного нерва в среднем мозге;
· псевдоуниполярные клетки, сгруппированные вблизи спинного мозга в межпозвоночных ганглиях;
· биполярные нейроны (имеют один аксон и один дендрит), расположенные в специализированных сенсорных органах — сетчатке глаза, обонятельном эпителии и луковице, слуховом и вестибулярном ганглиях;
· мультиполярные нейроны (имеют один аксон и несколько дендритов), преобладающие в ЦНС.
Развитие и рост нейрона[править | править вики-текст]

Конус роста
Нейрон развивается из небольшой клетки-предшественницы, которая перестаёт делиться ещё до того, как выпустит свои отростки. (Однако, вопрос о делении нейронов в настоящее время остаётся дискуссионным.) Как правило, первым начинает расти аксон, а дендриты образуются позже. На конце развивающегося отростка нервной клетки появляется утолщение неправильной формы, которое, видимо, и прокладывает путь через окружающую ткань. Это утолщение называется конусом роста нервной клетки. Он состоит из уплощенной части отростка нервной клетки с множеством тонкихшипиков. Микрошипики имеют толщину от 0,1 до 0,2 мкм и могут достигать 50 мкм в длину, широкая и плоская область конуса роста имеет ширину и длину около 5 мкм, хотя форма её может изменяться. Промежутки между микрошипиками конуса роста покрыты складчатой мембраной. Микрошипики находятся в постоянном движении — некоторые втягиваются в конус роста, другие удлиняются, отклоняются в разные стороны, прикасаются к субстрату и могут прилипать к нему.
Конус роста заполнен мелкими, иногда соединёнными друг с другом, мембранными пузырьками неправильной формы. Непосредственно под складчатыми участками мембраны и в шипиках находится плотная масса перепутанных актиновых филаментов. Конус роста содержит также митохондрии, микротрубочки и нейрофиламенты, аналогичные имеющимся в теле нейрона.
Вероятно, микротрубочки и нейрофиламенты удлиняются главным образом за счёт добавления вновь синтезированных субъединиц у основания отростка нейрона. Они продвигаются со скоростью около миллиметра в сутки, что соответствует скорости медленного аксонного транспорта в зрелом нейроне. Поскольку примерно такова и средняя скорость продвижения конуса роста, возможно, что во время роста отростка нейрона в его дальнем конце не происходит ни сборки, ни разрушения микротрубочек и нейрофиламентов. Новый мембранный материал добавляется, видимо, у окончания. Конус роста — это область быстрогоэкзоцитоза и эндоцитоза, о чём свидетельствует множество находящихся здесь пузырьков. Мелкие мембранные пузырьки переносятся по отростку нейрона от тела клетки к конусу роста с потоком быстрого аксонного транспорта. Мембранный материал, видимо, синтезируется в теле нейрона, переносится к конусу роста в виде пузырьков и включается здесь в плазматическую мембрану путём экзоцитоза, удлиняя таким образом отросток нервной клетки.
Росту аксонов и дендритов обычно предшествует фаза миграции нейронов, когда незрелые нейроны расселяются и находят себе постоянное место.
Нейроглия, или просто глия (от др.-греч. νεῦρον — волокно, нерв + γλία — клей), — совокупность вспомогательных клеток нервной ткани. Составляет около 40 % объёма ЦНС. Количество глиальных клеток в среднем в 10-50 раз больше, чем нейронов. Термин ввёл в 1846 году Рудольф Вирхов[1].
Глиальные клетки имеют общие функции и, частично, происхождение (исключение — микроглия). Они составляют специфическое микроокружение для нейронов, обеспечивая условия для генерации и передачи нервных импульсов, а также осуществляя часть метаболических процессов самого нейрона.
Нейроглия выполняет опорную, трофическую, секреторную, разграничительную и защитную функции.
11. Слуховой анализатор. Звукоулавливающий и звукопроводящий отделы слухового анализатора. Строение улитки, микроструктура кортиева органа. Механизм восприятия звуков разной частоты.
В Внутреннее ухо содержит рецепторный аппарат двух анализаторов: вестибулярного (преддверие и полукружные каналы) и слухового, к которому относится улитка с кортиевым органом.
Костная полость внутреннего уха, содержащая большое число камер и проходов между ними, называется лабиринтом. Он состоит из двух частей: костного лабиринта и перепончатого лабиринта. Костный лабиринт – это ряд полостей, расположенных в плотной части височной кости; в нем различают три составляющие: полукружные каналы – один из источников нервных импульсов, отражающих положение тела в пространстве; преддверие; и улитку – орган слуха.
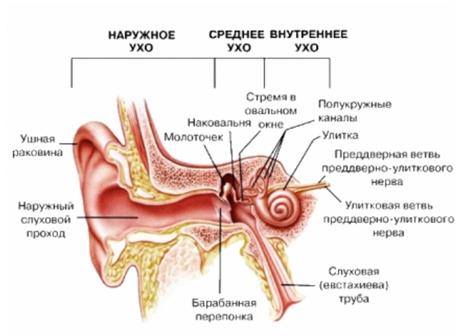
Строение уха
Перепончатый лабиринт заключен внутри костного лабиринта. Он наполнен жидкостью, эндолимфой, и окружен другой жидкостью – перилимфой, которая отделяет его от костного лабиринта. Перепончатый лабиринт, как и костный, состоит из трех основных частей. Первая соответствует по конфигурации трем полукружным каналам. Вторая делит костное преддверие на два отдела: маточку и мешочек. Удлиненная третья часть образует среднюю (улиточную) лестницу (спиральный канал), повторяющую изгибы улитки.
Полукружные каналы. Их всего шесть – по три в каждом ухе. Они имеют дугообразную форму и начинаются и кончаются в маточке. Три полукружных канала каждого уха расположены под прямыми углами друг к другу, один горизонтально, а два вертикально. Каждый канал имеет на одном конце расширение – ампулу. Шесть каналов расположены таким образом, что для каждого существует противолежащий ему канал в той же плоскости, но в другом ухе, однако их ампулы расположены на взаимнопротивоположных концах.
Улитка и кортиев орган. Название улитки определяется ее спирально извитой формой. Это костный канал, образующий два с половиной витка спирали и заполненный жидкостью. Завитки идут вокруг горизонтально лежащего стержня — веретена, вокруг которого наподобие винта закручена костная спиральная пластинка, пронизанная тонкими канальцами, где проходят волокна улитковой части преддверно-улиткового нерва — VIII пары черепно-мозговых нервов. Внутри, на одной стенке спирального канала по всей его длине расположен костный выступ. Две плоские мембраны идут от этого выступа к противоположной стенке так, что улитка по всей длине делится на три параллельных канала. Два наружных называются лестницей преддверия и барабанной лестницей, они сообщаются между собой у верхушки улитки. Центральный, т.н. спиральный, канал улитки, оканчивается слепо, а начало его сообщается с мешочком. Спиральный канал заполнен эндолимфой, лестница преддверия и барабанная лестница – перилимфой. Перилимфа имеет высокую концентрацию ионов натрия, тогда как эндолимфа – высокую концентрацию ионов калия. Важнейшей функцией эндолимфы, которая заряжена положительно по отношению к перилимфе, является создание на разделяющей их мембране электрического потенциала, обеспечивающего энергией процесс усиления входящих звуковых сигналов.
Лестница преддверия начинается в сферической полости – преддверии, лежащем в основании улитки. Один конец лестницы через овальное окно (окно преддверия) соприкасается с внутренней стенкой заполненной воздухом полости среднего уха. Барабанная лестница сообщается со средним ухом с помощью круглого окна (окна улитки). Жидкость
не может проходить через эти окна, так как овальное окно закрыто основанием стремени, а круглое – тонкой мембраной, отделяющей его от среднего уха. Спиральный канал улитки отделяется от барабанной лестницы т.н. основной (базилярной) мембраной, которая напоминает струнный инструмент в миниатюре. Она содержит ряд параллельных волокон различной длины и толщины, натянутых поперек спирального канала, причем волокна у основания спирального канала короткие и тонкие. Они постепенно удлиняются и утолщаются к концу улитки, как струны арфы. Мембрана покрыта рядами чувствительных, снабженных волосками клеток, составляющих т.н. кортиев орган, который выполняет высокоспециализированную функцию – превращает колебания основной мембраны в нервные импульсы. Волосковые клетки связаны с окончаниями нервных волокон, по выходе из кортиева органа образующих слуховой нерв (улитковую ветвь преддверно-улиткового нерва).
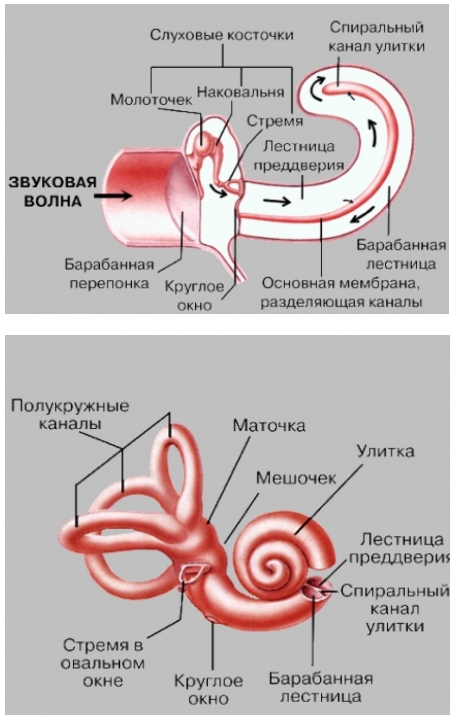
Улитка уха
Перепончатый улитковый лабиринт, или проток, имеет вид слепого преддверного выпячивания, находящегося в костной улитке и слепо заканчивающегося на ее верхушке. Он заполнен эндолимфой и представляет собой соединительно-тканный мешок длиной около35 мм. Улитковый проток разделяет костный спиральный канал на три части, занимая среднюю из них — средняя лестница (scalamedia), или улитковый ход, или улиточный канал. Верхняя часть — это лестница преддверия (scalavestibuli), или вестибулярная лестница, нижняя — барабанная, или тимпанальная, лестница (scalatympani). В них находится пери-лимфа. В области купола улитки обе лестницы сообщаются между собой через отверстие улитки (геликотрему). Барабанная лестница простирается до основания улитки, где она заканчивается у круглого окна улитки, закрытого вторичной барабанной перепонкой. Лестница преддверия сообщается с перилимфатическим пространством преддверия. Следует отметить, что перилимфа по своему составу напоминает плазму крови и цереброспинальную жидкость; в ней преобладает содержание натрия. Эндолимфа отличается от перилимфы более высокой (в 100 раз) концентрацией ионов калия и более низкой (в 10 раз) концентрацией ионов натрия; по своему химическому составу она напоминает внутриклеточную жидкость. По отношению к пери-лимфе она заряжена положительно.
Улитковый проток на поперечном разрезе имеет треугольную форму. Верхняя — преддверная стенка улиткового протока, обращенная к лестнице преддверия, образована тонкой преддверной (рейсснеровой) мембраной (membranavestibularis), которая изнутри покрыта однослойным плоским эпителием, а снаружи — эндотелием. Между ними расположена тонкофибриллярная соединительная ткань. Наружная стенка срастается с надкостницей наружной стенки костной улитки и представлена спиральной связкой, которая имеется во всех завитках улитки. На связке расположена сосудистая полоска (striavascularis), богатая капиллярами и покрытая кубическими клетками, которые продуцируют эндолимфу. Нижняя — барабанная стенка, обращенная к барабанной лестнице, устроена наиболее сложно. Она представлена базилярной мембраной, или пластинкой (laminabasilaris), на которой располагается спиральный, или кортиев орган, осуществляющий восприятие звуков. Плотная и упругая базиляр-ная пластинка, или основная мембрана, одним концом прикрепляется к спиральной костной пластинке, противоположным — к спиральной связке. Мембрана образована тонкими слабо натянутыми радиальными коллагеновыми волокнами (около 24 тыс.), длина которых возрастает от основания улитки к ее вершине — вблизи овального окна ширина базилярной мембраны составляет0,04 мм, а затем по направлению к вершине улитки, постепенно расширяясь, она достигает в конце0,5 мм(т.е. базилярная мембрана расширяется там, где улитка сужается). Волокна состоят из тонких анастомозирующих между собой фибрилл. Слабое натяжение волокон базилярной мембраны создает условия для их колебательных движений.
Собственно орган слуха — кортиев орган — находится в костной улитке. Кортиев орган — рецепторная часть слухового анализатора, расположенная внутри перепончатого лабиринта. В процессе эволюции возникает на основе структур боковых органов. Воспринимает колебания волокон, расположенных в канале внутреннего уха, и передаёт в слуховую зону коры больших полушарий, где и формируются звуковые сигналы. В Кортиевом органе начинается первичное формирование анализа звуковых сигналов.
Расположение. Кортиев орган располагается в спирально завитом костном канале внутреннего уха — улитковом ходе, заполненном эндолимфой и перилимфой. Верхняя стенка хода прилегает к т. н. лестнице преддверия и называется рейснеровой перепонкой; нижняя стенка, граничащая с т. н. барабанной лестницей, образована основной перепонкой, прикрепляющейся к спиральной костной пластинке. Кортиев орган представлен опорными, или поддерживающими, клетками, и рецепторными клетками, или фонорецепторами. Выделяют два типа опорных и два типа рецепторных клеток — наружные и внутренние.
Наружные опорные клетки лежат дальше от края спиральной костной пластинки, а внутренние — ближе к нему. Оба вида опорных клеток сходятся под острым углом друг к другу и образуют канал треугольной формы — внутренний (кортиев) туннель, заполненный эндо-лимфой, который проходит спирально вдоль всего корти-ева органа. В туннеле расположены безмиелиновые нервные волокна, идущие от нейронов спирального ганглия.
Фонорецепторы лежат на опорных клетках. Они представляют собой вторично-чувствующие рецепторы (механорецепторы), трансформирующие механические колебания в электрические потенциалы. Фонорецепторы (на основании их отношения к кортиеву туннелю) подразделяются на внутренние (колбообразной формы) и наружные (цилиндрической формы), которые отделены друг от друга кортиевыми дугами. Внутренние волосковые клетки располагаются в один ряд; их общее число по всей длине перепончатого канала достигает 3500. Наружные волосковые клетки располагаются в 3-4 ряда; их общее число достигает 12000-20000. Каждая волосковая клетка имеет удлиненную форму; один ее полюс приближен к основной мембране, второй находится в полости перепончатого канала улитки. На конце этого полюса есть волоски, или стереоцилии (до 100 в клетке). Волоски рецепторных клеток омываются эндолимфой и контактируют с покровной, или текториальной, мембраной (membranatectoria), которая по всему ходу перепончатого канала расположена над волосковыми клетками. Эта мембрана имеет желеобразную консистенцию, один край которой прикрепляется к костной спиральной пластинке, а другой свободно оканчивается в полости улиткового протока чуть дальше наружных рецепторных клеток.
Все фонорецепторы, независимо от локализации, синаптически связаны с 32000 дендритов биполярных чувствительных клеток, находящихся в спиральном нервном ганглии улитки. Эти первыенейроны слухового пути, аксоны которых образуют улитковую (кохлеарную) часть VIII пары черепно-мозговых нервов; они передают сигналы на кохлеарные ядра продолговатого мозга. При этом сигналы от каждой внутренней волосковои клетки передаются на биполярные клетки одновременно по нескольким волокнам (вероятно, это повышает надежность передачи информации), в то время как сигналы от нескольких наружных волосковых клеток конвергируют на одном волокне. Поэтому около 95% волокон слухового нерва несет информацию в продолговатый мозг от внутренних волосковых клеток (хотя их количество не превышает 3500), а 5% волокон передают информацию от наружных волосковых клеток, число которых достигает 12000-20000. Эти данные подчеркивают огромную физиологическую значимость внутренних волосковых клеток в рецепции звуков.
К волосковым клеткам подходят и эфферентные волокна — аксоны нейронов верхней оливы. Волокна, приходящие к внутренним волосковым клеткам, оканчиваются не на самих этих клетках, а на афферентных волокнах. Предполагается, что они оказывают тормозное воздействие на передачу слухового сигнала, способствуя обострению частотного разрешения. Волокна, приходящие к наружным волосковым клеткам, воздействуют на них непосредственно и за счет изменения их длины, меняют их фоночувствительность. Таким образом, с помощью эфферентных оливо-кохлеарных волокон (волокон пучка Расмуссена) высшие акустические центры регулируют чувствительность фонорецепторов и поток афферентных импульсов от них к мозговым центрам.
Проведение звуковых колебаний в улитке. Восприятие звука осуществляется с участием фонорецепторов. Их возбуждение под влиянием звуковой волны приводит к генерации рецепторного потенциала, который вызывает возбуждение дендритов биполярногонейрона спирального ганглия. Но каким образом осуществляется кодирование частоты и силы звука? Это один из наиболее сложных вопросов физиологии слухового анализатора.
Современное представление о кодировании частоты и силы звука сводится к следующему. Звуковая волна, воздействуя на систему слуховых косточек среднего уха, приводит в колебательное движение мембрану овального окна преддверия, которая, прогибаясь, вызывает волнообразные перемещения перилимфы верхнего и нижнего каналов, которые постепенно затухают по направлению к вершине улитки. Поскольку все жидкости несжимаемы, колебания эти были бы невозможны, если бы не мембрана круглого окна, которая выпячивается при надавливании основания стремечка на овальное окно и принимает исходное положение при прекращении давления. Колебания перилимфы передаются на вестибулярную мембрану, а также на полость среднего канала, приводя в движение эндолимфу и базиляр-ную мембрану (вестибулярная мембрана очень тонкая, поэтому жидкость в верхнем и среднем каналах колеблется так, как будто оба канала едины). При действии на ухо звуков низкой частоты (до 1000 Гц) происходит смещение базилярной мембраны на всем ее протяжении от основания до верхушки улитки. При увеличении частоты звукового сигнала происходит перемещение укороченного по длине колеблющегося столба жидкости ближе к овальному окну, к наиболее жесткому и упругому участку базилярной мембраны. Деформируясь, базилярная мембрана смещает волоски волосковых клеток относительно текториальной мембраны. В результате такого смещения возникает электрический разряд волосковых клеток. Существует прямая зависимость между амплитудой смещения основной мембраны и количеством вовлекаемых в процесс возбуждения нейронов слуховой коры.
| Механизм проведения звуковых колебаний в улитке Звуковые волны улавливаются ушной раковиной и через слуховой канал направляются к барабанной перепонке. Колебания барабанной перепонки, через систему слуховых косточек, передаются посредством стремечка мембране овального окна, и через нее передаются лимфатической жидкости. На колебания жидкости отзываются (резонируют), в зависимости от частоты колебаний, только определенные волокна главной мембраны. Волосковые клетки Кортиева органа возбуждаются от прикосновения к ним волокон главной мембраны и по слуховому нерву передаются в мозгимпульсы, где и создается окончательное ощущение звука. http://www.braintools.ru/article/9775 |
Пути проведения звука. С функциональной точки зрения орган слуха (периферическая часть слухового анализатора) делится на две части:
1. звукопроводящий аппарат - наружное и среднее ухо, а также некоторые элементы (перилимфа и эндолимфа) внутреннего уха;
2. звуковоспринимающий аппарат - внутреннее ухо.
12. Синапсы, их классификация, строение. Механизм проведения возбуждения через возбуждающие и тормозные синапсы. Свойства синапсов.
Си́напс [1] (греч. σύναψις, от συνάπτειν — обнимать, обхватывать, пожимать руку) — место контакта между двумя нейронами или между нейроном и получающей сигнал эффекторной клеткой. Служит для передачи нервного импульса между двумя клетками, причём в ходе синаптической передачи амплитуда и частота сигнала могут регулироваться. Передача импульсов осуществляется химическим путём с помощью медиаторов или электрическим путём посредством прохождения ионов из одной клетки в другую.
Термин был введён в 1897 г. английским физиологом Чарльзом Шеррингтоном. Однако сам Шеррингтон утверждал, что получил идею этого термина в разговоре от физиолога Майкла Фостера[2].
Содержание
[убрать]
· 1 Классификации синапсов
o 1.1 По механизму передачи нервного импульса
§ 1.1.1 По местоположению и принадлежности структурам
§ 1.1.2 По нейромедиатору
o 1.2 По знаку действия
· 2 Механизм функционирования химического синапса
· 3 История открытия
· 4 См. также
· 5 Примечания
· 6 Ссылки
Классификации синапсов[править | править вики-текст]

Основные элементы электрического синапса (эфапса): а — коннексон в закрытом состоянии; b — коннексон в открытом состоянии; с — коннексон, встроенный в мембрану; d — мономер коннексина, е —плазматическая мембрана; f — межклеточное пространство; g — промежуток в 2-4 нанометра в электрическом синапсе; h — гидрофильный канал коннексона.
По механизму передачи нервного импульса[править | править вики-текст]
· химический — это место близкого прилегания двух нервных клеток, для передачи нервного импульса через которое клетка-источник выпускает в межклеточное пространство особое вещество, нейромедиатор, присутствие которого в синаптической щели возбуждает или затормаживает клетку-приёмник.
· электрический (эфапс) — место более близкого прилегания пары клеток, где их мембраны соединяются с помощью особых белковых образований — коннексонов (каждый коннексон состоит из шести белковых субъединиц). Расстояние между мембранами клетки в электрическом синапсе — 3,5 нм (обычное межклеточное — 20 нм). Так как сопротивление внеклеточной жидкости мало(в данном случае), импульсы через синапс проходят не задерживаясь. Электрические синапсы обычно бывают возбуждающими.
· смешанные синапсы — Пресинаптический потенциал действия создает ток, который деполяризует постсинаптическую мембрану типичного химического синапса, где пре- и постсинаптические мембраны не плотно прилегают друг к другу. Таким образом, в этих синапсах химическая передача служит необходимым усиливающим механизмом.
Наиболее распространены химические синапсы. Для нервной системы млекопитающих электрические синапсы менее характерны, чем химические.
По знаку действия[править | править вики-текст]
· возбуждающие
· тормозные.
Если первые способствуют возникновению возбуждения в постсинаптической клетке (в них в результате поступления импульса происходит деполяризация мембраны, которая может вызвать потенциал действия при определённых условиях.), то вторые, напротив, прекращают или предотвращают его появление, препятствуют дальнейшему распространению импульса. Обычно тормозными являются глицинергические (медиатор — глицин) и ГАМК-ергические синапсы (медиатор — гамма-аминомасляная кислота).
Тормозные синапсы бывают двух видов: 1) синапс, в пресинаптических окончаниях которого выделяется медиатор, гиперполяризующий постсинаптическую мембрану и вызывающий возникновение тормозного постсинаптического потенциала; 2) аксо-аксональный синапс, обеспечивающий пресинаптическое торможение. Синапс холинергический (s. cholinergica) — синапс, медиатором в котором является ацетилхолин.
В некоторых синапсах присутствует постсинаптическое уплотнение — электронно-плотная зона, состоящая из белков. По её наличию или отсутствию выделяют синапсы асимметричные и симметричные. Известно, что все глутаматергические синапсы асимметричны, а ГАМКергические — симметричны.
В случаях, когда с постсинаптической мембраной контактирует несколько синаптических расширений, образуются множественные синапсы.
К специальным формам синапсов относятся шипиковые аппараты, в которых с синаптическим расширением контактируют короткие одиночные или множественные выпячивания постсинаптической мембраны дендрита. Шипиковые аппараты значительно увеличивают количество синаптических контактов на нейроне и, следовательно, количество перерабатываемой информации. «Не-шипиковые» синапсы называются «сидячими». Например, сидячими являются все ГАМК-ергические синапсы.
Механизм функционирования химического синапса[править | править вики-текст]
Основная статья: Химический синапс
Типичный синапс — аксо-дендритический химический. Такой синапс состоит из двух частей: пресинаптической, образованной булавовидным расширением окончанием аксона передающей клетки и постсинаптической, представленной контактирующим участком плазматической мембраны воспринимающей клетки (в данном случае — участком дендрита).
Между обеими частями имеется синаптическая щель — промежуток шириной 10—50 нм между постсинаптической и пресинаптической мембранами, края которой укреплены межклеточными контактами.
Часть аксолеммы булавовидного расширения, прилежащая к синаптической щели, называется пресинаптической мембраной. Участок цитолеммы воспринимающей клетки, ограничивающий синаптическую щель с противоположной стороны, называется постсинаптической мембраной, в химических синапсах она рельефна и содержит многочисленные рецепторы.
В синаптическом расширении имеются мелкие везикулы, так называемые синаптические пузырьки, содержащие либо медиатор (вещество-посредник в передаче возбуждения), либо фермент, разрушающий этот медиатор. На постсинаптической, а часто и на пресинаптической мембранах присутствуют рецепторы к тому или иному медиатору.
При деполяризации пресинаптическойтерминали открываются потенциал-чувствительные кальциевые каналы, ионы кальция входят в пресинаптическуютерминаль и запускают механизм слияния синаптических пузырьков с мембраной. В результате медиатор выходит в синаптическую щель и присоединяется к белкам-рецепторам постсинаптической мембраны, которые делятся на метаботропные и ионотропные. Первые связаны с G-белком и запускают каскад реакций внутриклеточной передачи сигнала. Вторые связаны с ионными каналами, которые открываются при связывании с ними нейромедиатора, что приводит к изменению мембранного потенциала. Медиатор действует в течение очень короткого времени, после чего разрушается специфическим ферментом. Например, в холинэргических синапсах фермент, разрушающий медиатор в синаптической щели — ацетилхолинэстераза. Одновременно часть медиатора может перемещаться с помощью белков-переносчиков через постсинаптическую мембрану (прямой захват) и в обратном направлении через пресинаптическую мембрану (обратный захват). В ряде случаев медиатор также поглощается соседними клетками нейроглии.
Открыты два механизма высвобождения: с полным слиянием везикулы с плазмалеммой и так называемый «поцеловал и убежал» (англ. kiss-and-run), когда везикула соединяется с мембраной, и из неё в синаптическую щель выходят небольшие молекулы, а крупные остаются в везикуле. Второй механизм, предположительно, быстрее первого, с помощью него происходит синаптическая передача при высоком содержании ионов кальция в синаптической бляшке.
Следствием такой структуры синапса является одностороннее проведение нервного импульса. Существует так называемая синаптическая задержка — время, нужное для передачи нервного импульса. Её длительность составляет около — 0,5 мс.
Так называемый «принцип Дейла» (один нейрон — один медиатор) признан ошибочным. Или, как иногда считают, он уточнён: из одного окончания клетки может выделяться не один, а несколько медиаторов, причём их набор постоянен для данной клетки.
13. Эритроциты, их количество, строение, функции, образование. Гемоглобин, его количество, строение, соединения гемоглобина.
Эритроци́ты (от греч. ἐρυθρός — красный и κύτος — вместилище, клетка), также известные под названием кра́сныекровяны́е тельца́, — постклеточные структуры крови позвоночных животных (включая человека) и гемолимфы некоторых беспозвоночных (сипункулид, у которых эритроциты плавают в полости целома[1] и некоторых двустворчатых моллюсков[2]). Они насыщаются кислородом в лёгких или в жабрах и затем разносят его по телу животного.
Их цитоплазма богата гемоглобином — пигментом красного цвета, содержащим атом железа, который способен связывать кислород и придаёт эритроцитам красный цвет.
Человеческие эритроциты — очень маленькие эластичные клетки дисковидной двояковогнутой формы диаметром от 7 до 10 мкм. Размер и эластичность способствуют им при движении по капиллярам, их форма повышает площадь поверхности и облегчает газообмен. В них отсутствует клеточное ядро и большинство органелл, что повышает содержание гемоглобина. Около 2,4 миллиона новых эритроцитов образуется в костном мозге каждую секунду[3]. Они циркулируют в крови около 100—120 дней и затем поглощаются макрофагами. Приблизительно четверть всех клеток в теле человека — эритроциты[4].
Структура и состав[править | править

