 2015-01-07
2015-01-07 10714
10714Все комплексы дыхательной цепи интегрированы во внутренней мембране митохондрий (рис. 21). Протон генерирующими являются комплексы I, III и IV.
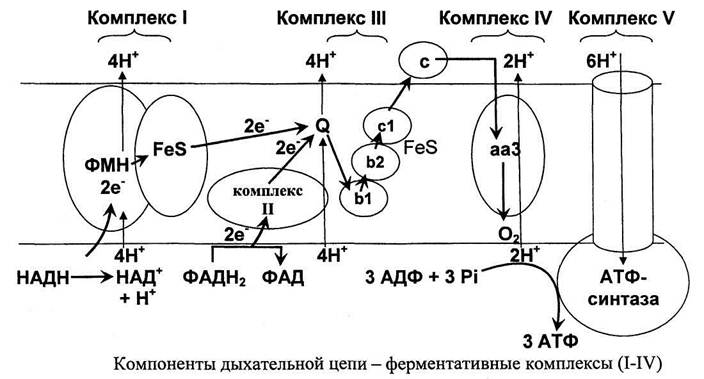
Электроны поступают в дыхательную цепь различными путями. При окислении НАДН в комплекс I происходит перенос 2-х электронов от ФМН и FeS-центров на убихинон (Q). При этом через комплекс I переносятся 4 Н+.
Образующиеся при окислении сукцината, ацетил-КоА и других субстратов электроны переносятся на убихинон (Q) комплексом II через связанный с ферментом ФАДНг или через электрон переносящий флавопротеин (EFT). Прохождение пары электронов через комплекс II сопровождается переносом 4-х протонов из матрик-са.
Последний протон генерирующий комплекс (комплекс IV) - фермент цитохром С оксидаза обеспечивает образование воды из электронов поступающих в него по дыхательной цепи, кислорода и протонов митохондриального матрикса. При этом через комплекс IV переносятся 2 Н+.
Таким образом, миграция пары электронов по полной дыхательной цепи сопровождается выбросом 10 протонов из матрикса. Создается трансмембранный электрохимический градиент, обусловленный различной концентрацией протонов в матриксе и межмембранном пространстве. Энергия этого электрохимического градиента расходуется на синтез АТФ в V комплексе дыхательной цепи (фермент - протон-транслоцирующая АТФ синтаза). Синтез АТФ сопряжен с обратным потоком протонов в матрикс (рис. 22).
Трансмембранный электрохимический потенциал как промежуточная форма энергии при окислительном фосфорилировании
Преобразование энергии электронов в энергию АТФ происходит в два этапа. На первом энергия электронов преобразуется в энергию протонного градиента. Концентрация протонов в межмембранном пространстве больше чем в матриксе митохондрии. Поскольку каждый протон несет положительный заряд, вследствие этого появляется разность потенциалов по обе стороны мембраны. В совокупности электрический и концентрационный градиенты составляют электрохимический потенциал (ДцН4). АцН* - Источник энергии для синтеза АТФ. Активный транспорт протонов происходит в трех участках дыхательной цепи, в ферментативных комплексах I, III и IV (рис. 21). Эти участки часто называют пунктами сопряжения дыхания и фосфорилирования, хотя правильнее считать пунктом сопряжения окисления (дыхания) и фосфорилирования комплекс V, в котором синтезируется АТФ.
На втором этапе синтезируется АТФ.
Синтез АТФ. Фермент КГ-транслоцирующая АТФ синтаза состоит из двух крупных белковых комплексов, каждый из которых построен из многих субъединиц. В структуре этого фермента выделяют протонный канал (Fo) и каталитическую часть фермента (Fi) содержащую три активных центра. В каждый из активных-центров поступают молекулы АДФ и неорганического фосфата (Pi) Протонный канал пронизывает внутреннюю митохондриальную мембрану, а каталитическая часть фермента представляет собой крупное грибовидное образование, обращенное в сторону матрикса. Ультразвуковая обработка интактных (неповрежденных) митохондрий вызывает остановку синтеза АТФ, поскольку каталитическая часть фермента Fi отделяется от протонного канала. При этом поступление потока протонов из межмембранного пространства в митохондриальныи матрикс не нарушается.
Под действием потока протонов по каналу каталитическая часть АТФ синтазы (Fo) вращается вокруг устья канала. Синтез АТФ проходит три стадии:
1) поступление АДФ и неорганического фосфата (Р;) в каталитический центр,
2) поворот на 120° и сжимающая деформация каталитического центра, сближаю
щая отталкивающиеся группы остатков фосфорной кислоты; на этой стадии про
исходит образование АТФ и молекулы воды,
3) поворот на 120° и освобождение из активного центра молекулы АТФ.
Затем следует поворот на 120° АТФ синтазы и в освободившийся каталитический центр поступают новые молекулы АДФ и Pi (неорганического фосфата)

