 2015-04-30
2015-04-30 1140
1140Структура и функции желчных кислот. Основными эмульгаторами ли пидов в тонком кишечнике являются желчные кислоты, содержащиеся в виде натриевых солей в желчи, поступающей в двенадцатиперстную кишку из желчного пузыря. В желчи содержится ряд различных желчных кислот, кото рые в печени образуются из холестерола. По химическому строению желчные кислоты являются производными холановой кислоты и отличаются друг от друга числом и положением гидроксигрупп:
Первым этапом этого процесса является гидроксилирование холестерола под действием фермента 7а-гидроксилазы с участием кислорода, НАДФН и цитохрома. Это основная регуляторная реакция синтеза желчных кислот: фермент активируется витамином С и ингибируется по типу обратной связи желчной кислотой. На этой стадии идет разделение путей синтеза желчных кислот: одна ветвь ведет к синтезу холевой кислоты, другая — к образованию хенодезоксихолевой кислоты. Реакции синтеза обеих кислот включают гидроксилирование и укорачивание боковой цепи.
Кроме роли эмульгаторов, желчные кислоты в кишечнике способствуют всасыванию продуктов расщепления жиров и активируют панкреатическую липазу. ■ Кроме желчных кислот, соли жирных кислот в комплексе с моноаии -1 - церолами и ненасыщенными жирными кислотами, будучи поверхностно- активными веществами, также способствуют эмульгированию жира и стабилизации образовавшейся эмульсии.
Панкреатическая липаза— это гликопротеин, секретируется поджелудочной железой в виде неактивно предшественника — пролипазы, В верхней части кишечника происходит активация пролипазы путем образования комплекса с низкомолекулярным белком — колипазой. Известно активирующее и стабилизирующее действие желчных кислот на панкреатическую липазу, хотя механизм его остается неясным.
Специфичность действия липазы определяется положением эфирных связей в триацилглицероле.
45. Окисление жирных кислот (β-окисление). ПОСЛЕДОВАТЕЛЬНОСТЬ. РЕАКЦИИ
протекает в печени, почках, скелетных и сердечной мышцах, в жировой ткани. В мозговой ткани скорость окисления жирных кислот весьма незначительна; основным источником энергии в мозговой ткани служит глюкоза.
окисление молекулы жирной кислоты в тканях организма происходит в β-положении. В результате от молекулы жирной кислоты последовательно отщепляются двууглеродные фрагменты со стороны карбоксильной группы.
Жирные кислоты, входящие в состав естественных жиров животных и растений, имеют четное число углеродных атомов. Любая такая кислота, от которой отщепляется по паре углеродных атомов, в конце концов проходит через стадию масляной кислоты. После очередного β-окисления масляная кислота становится ацетоуксусной. Последняя затем гидролизуется до двух молекул уксусной кислоты.
Доставка жирных кислот к месту их окисления – к митохондриям – происходит сложным путем: при участии альбумина осуществляется транспорт жирной кислоты в клетку; при участии специальных белков (fatty acid binding proteins, FABP) – транспорт в пределах цитозоля; при участии карнитина – транспорт жирной кислоты из цитозоля в митохондрии.
Процесс окисления жирных кислот складывается из следующих основных этапов.
Активация жирных кислот. Свободная жирная кислота независимо от длины углеводородной цепи является метаболически инертной и не может подвергаться никаким биохимическим превращениям, в том числе окислению, пока не будет активирована. Активация жирной кислоты протекает на наружной поверхности мембраны митохондрий при участии АТФ, коэнзима A (HS-KoA) и ионов Mg2+. Реакция катализируется ферментом ацил-КоА-синтетазой:
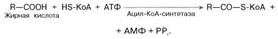
В результате реакции образуется ацил-КоА, являющийся активной формой жирной кислоты.
Считают, что активация жирной кислоты протекает в 2 этапа. Сначала жирная кислота реагирует с АТФ с образованием ациладенилата, представляющим собой эфир жирной кислоты и АМФ. Далее сульфгидрильная группа КоА действует на прочно связанный с ферментом ациладенилат с образованием ацил-КоА и АМФ.
Транспорт жирных кислот внутрь митохондрий. Коэнзимная форма жирной кислоты, в равной мере как и свободные жирные кислоты, не обладает способностью проникать внутрь митохондрий, где, собственно, и протекает их окисление. Переносчиком активированных жирных кислот с длинной цепью через внутреннюю митохондриальную мембрану служит карнитин. Ацильная группа переносится с атома серы КоА на гидроксильную группу карнитина с образованием ацилкарнитина, который диффундирует через внутреннюю митохондриальную мембрану:
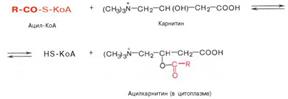
Реакция протекает при участии специфического цитоплазматического фермента карнитин-ацилтрансферазы. Уже на той стороне мембраны, которая обращена к матриксу, ацильная группа переносится обратно на КоА, что термодинамически выгодно, поскольку О-ацильная связь в кар-нитине обладает высоким потенциалом переноса группы. Иными словами, после прохождения ацилкарнитина через мембрану митохондрий происходит обратная реакция – расщепление ацилкарнитина при участии HS-KoA и митохондриальной карнитин-ацилтрансферазы:
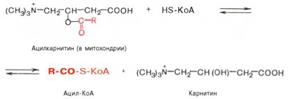
Внутримитохондриальное окислениежирных кислот. Процесс окисления жирной кислоты в митохондриях клетки включает несколько последовательных энзиматических реакций.
Первая стадия дегидрирования. Ацил-КоА в митохондриях прежде всего подвергается ферментативному дегидрированию, при этом ацил-КоА теряет 2 атома водорода в α- и β-положениях, превращаясь в КоА-эфир ненасыщенной кислоты. Таким образом, первой реакцией в каждом цикле распада ацил-КоА является его окисление ацил-КоА-де-гидрогеназой, приводящее к образованию еноил-КоА с двойной связью между С-2 и С-3:
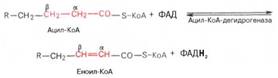
Существует несколько ФАД-содержащих ацил-КоА-дегидрогеназ, каждая из которых обладает специфичностью по отношению к ацил-КоА с определенной длиной углеродной цепи.
Стадия гидратации. Ненасыщенный ацил-КоА (еноил-КоА) при участии фермента еноил-КоА-гидратазы присоединяет молекулу воды. В результате образуется β-оксиацил-КоА (или 3-гидроксиацил-КоА):
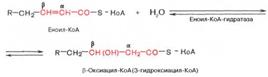
Заметим, что гидратация еноил-КоА стереоспецифична, подобно гидратации фумарата и аконитата (см. с. 348). В результате гидратации транс-Δ2-двойной связи образуется только L-изомер 3-гидроксиацил-КоА.
Вторая стадия дегидрирования. Образовавшийся β-оксиацил-КоА (3-гидроксиацил-КоА) затем дегидрируется. Эту реакцию катализируют НАД+-зависимые дегидрогеназы:
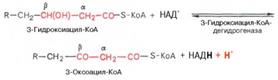
Тиолазная реакция. В ходе предыдущих реакций происходило окисление метиленовой группы при С-3 в оксогруппу. Тиолазная реакция представляет собой расщепление 3-оксоацил-КоА с помощью тиоловой группы второй молекулы КоА. В результате образуется укороченный на два углеродных атома ацил-КоА и двууглеродный фрагмент в виде ацетил-КоА. Данная реакция катализируется ацетил-КоА-ацилтрансферазой (β-ке-тотиолазой):
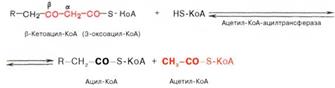
Образовавшийся ацетил-КоА подвергается окислению в цикле трикар-боновых кислот, а ацил-КоА, укоротившийся на два углеродных атома, снова многократно проходит весь путь β-окисления вплоть до образования бутирил-КоА (4-углеродное соединение), который в свою очередь окисляется до 2 молекул ацетил-КоА
За один цикл β-окисления образуется 1 молекула ацетил-СоА, окисление которого в цитратном цикле обеспечивает синтез 12 моль ATP. Кроме того, образуется 1 моль FADH2 и 1 моль NADH+H, при окислении которых в дыхательной цепи синтезируется соответственно 2 и 3 моль ATP (в сумме 5).
Таким образом, при окислении, например, пальмитиновой кислоты (С16) происходит 7 циклов β-окисления, в результате которых образуется 8 моль ацетил-СоА, 7 моль FADH2 и 7 моль NADH+H. Следовательно, выход ATP составляет 35 молекул в результате β-окисления и 96 ATP в результате цитратного цикла, что соответствует в сумме 131 молекул АТФ.
Суммарное уравнение β-окисления пальмитиновой кислоты
Пальмитоил-КоА + 7ФАД + 7НАД+ + 7Н2O + 7HS-KoA –>
–> 8Ацетил-КоА + 7ФАДН2 + 7НАДН + 7Н+.
Изменение свободной энергии ΔF при полном сгорании 1 моля пальмитиновой кислоты составляет 2338 ккал, а богатая энергией фосфатная связь ATP характеризуется величиной 7,6 ккал/моль. Нетрудно подсчитать, что примерно 990 ккал (7,6 х 130), или 42% от всей потенциальной энергии пальмитиновой кислоты при ее окислении в организме, используется для ресинтеза АТР, а оставшаяся часть теряется в виде тепла.
Окисление ненасыщенных жирных кислот в принципе происходит так же, как и окисление насыщенных жирных кислот, но с некоторыми особенностями. Двойные связи природных ненасыщенных жирных кислот (олеиновой, линолевой и т.д.) имеют цис-конфигурацию, а в КоА-эфирах ненасыщенных кислот, являющихся промежуточными продуктами при β-окислении насыщенных жирных кислот, двойные связи имеют трансконфигурацию. Кроме того, последовательное удаление двууглеродных фрагментов при окислении ненасыщенных жирных кислот до первой двойной связи дает Δ3,4-ацил-КоА, а не Δ2,3-ацил-КоА, который является промежуточным продуктом при β-окислении ненасыщенных жирных кислот:
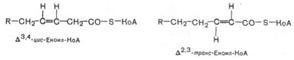
В тканях существует фермент, который осуществляет перемещение двойной связи из положения 3–4 в положение 2–3, а также изменяет конфигурацию двойной связи из цис- в транс-положение. Этот фермент получил название Δ3,4-цис –> Δ2,3-транс-еноил-КоА-изомеразы. На рис. 11.3 представлен путь β-окисления олеиновой кислоты, иллюстрирующий назначение данного фермента
Окисление жирных кислот с нечетным числом углеродных атомов. Как отмечалось, основная масса природных липидов содержит жирные кислоты с четным числом углеродных атомов. Однако в липидах многих растений и некоторых морских организмов присутствуют жирные кислоты с нечетным числом атомов углерода. Кроме того, у жвачных животных при переваривании углеводов в рубце образуется большое количество про-пионовой кислоты, которая содержит три углеродных атома. Пропионат всасывается в кровь и окисляется в печени и других тканях. Установлено, что жирные кислоты с нечетным числом углеродных атомов окисляются таким же образом, как и жирные кислоты с четным числом углеродных атомов, с той лишь разницей, что на последнем этапе расщепления (β-окисления) образуется одна молекула пропионил-КоА и одна молекула ацетил-КоА, а не 2 молекулы ацетил-КоА.
46. СИНТЕЗ ЖИРОВ ИЗ УГЛЕВОДОВ В ПЕЧЕНИ И ЖИРОВОЙ ТКАНИ. ВЛИЯНИЕ ИНСУЛИНА, ОЖИРЕНИЕ
Синтез жиров происходит в абсорбтивный период в печени и жировой ткани. Непосредственными субстратами в синтезе жиров являются ацил-КоА и глицерол-3-фосфат.

