 2015-05-14
2015-05-14 3832
38321896 жылы Н. В. Гамалея кейбір бактериялар сүзіндісінің басқа бактериялардың клеткаларын ерітіп (лизис) жіберетін қасиетін анықтады.Ол топалаң вирусын дистильденген ‘суда ерітіп, оны жаңадан өсірілген топалаң вирусына жұқтырады. Сонда бұл ерітінді жаңадан өсірілген вирусты ерітіп, қүртып жіберген. Мұны бактериофагтар деп атады. Дәлме-дәл аударғанда ол бактерия-ларды «жеушілер» немесе «жалмаушылар» деген сөз.
Бүдан кейінгі зерттеулер басқа бактерияларды ерітіп жіберетін, табиғатта толып жатқан бактериофагтардың барлығын анықтады. Бактериофагтардың жеке түрлері белгілі бір бактерияны зақымдай алады. Ол тек ауру қоздырғыш бактерияларды ғана жойып жібермей, пайдалы түрлерін де жоя алады.
Бактериофагтардың пішіні де алуан түрлі. Олардың кейбіреулері шар тәрізді қосалқы бүтақшалары бар денелер. Кейбір түрлері итбалыққа ұқсайды. Өз бұтақшасымен фагтар бактерия клеткасына бекиді. Олар бактерия клеткаларының іқабығын ерітетін лтізоцим дөп аталатын, фермент бөліп шығарады. Еріген жерге фагтың ішіндегі заттар бактердя клеткасына «енеді де, оның бұтақшасы мен сыртқы қабығы бактерия клеткасының сыртында қалып қояды. Фаг клетка ішіне кірісімен өзінің зиянды әрекетін ти-гізе бастайды. Бактерия клеткасы бұдан кейін ісініп, жарылады да, оданчжаңадан дамыған фаг бөлінга шығып, ол келесі клетканы заікыімдаға кіріседі.
Фагтыіі көбею стадиясының ұзақтығы 30-дан 90 минутқа дейін созылады.
Бактериофагтардың шамасы 40—100 мк-ға тең.
Олардың бактериялар сияқты клеткалық құрылысы болмайды және тек тірі клеткада тіршілік етуге бейімделген. Ол құрғақшылыққа, төменгі температураға, көптеген химиялық уларға төзімді. Бірақ 50%-тік глицерин ерітіндісі бактериофагтарды қырып жібере-ді, ал +80°-қа дейінгі ыстықты олар оңай көтереді. Температура + 100°-қа көтерілгенде бактериофагтар қырылып кетеді. Фагтардың көбеюі небары 30—90 минутқа дейін созылады. Олар табиғатта өте көп тараған. Қазіргі кезде бактериофагтарды медицинада және ветеринарияда кейбір зиянды ауру коздырғыштарға карсы колда-нады.
3. Ген қызметінің бақылануы.
Ген(грек. genos — тұқым, тек) — тұқым қуалаудың қандай да бір элементар белгісін қалыптастыруға жауапты материалдық бірлік. Генде жасушаның құрылымы мен қызметін анықтайтын генетикалық ақпарат болады. Бір организмнің Гендер жиынтығы оның генотипін құрайды.Ген терминін алғаш рет 1909 жылы Дания ғалымы В.Йогансен енгізді. Барлық Гендер ДНҚ-дан тұрады және әрбір жеке жасушадағы мыңдаған осындай Гендер жеке ДНҚ молекуларының үзіндісі түрінде емес, хромосома деп аталатын, ірі құрылымдық бірлік құрамында болады. Жасушаның бөлінуі кезінде бұл хромосомалар екі еселенеді және жаңа түзілген жас жасушаалар осындай ата-аналық Гендер жиынтығының көшірмесін алады. Соның нәтижесінде жасушааның барлық белгілері (қасиеттері) ұрпақтан ұрпаққа беріледі, яғни тұқым қуалайды. Әртүрлі органимздердегі Геннің орташа ұзындығы 1000 нуклеотид негіздерінің жұбынан құралады деп есептеуге болады, Гені, тағыда басқа Гаметалардың түзілуі мен ұрықтану процестеріндегі әртүрлі Гендер бойынша белгілердің тәуелсіз ажырауы мен гомологтық емес хромосомалар әрекетінің арасындағы қатарластық (параллелизм), тұқым қуалаушылықтың хромосомалық теориясының негізін қалады. Бұл теория бойынша Гендер хромосомаларда тізбектеле орналасады да, олар тұқым қуалаушылықтың материалдық негізін қалайды (қ. Мейоз). Жасушадағы ақуыздың синтезделуі және олардың қарым-қатынасы туралы ақпарат тек Гендерде болады, яғни әрбір Ген белгілі бір ақуыз (полипептидті тізбек) синтезіне жауапты. Ақуыз синтезін бақылай отырып, Ген организмдегі барлық химиялық реакцияларды басқарады, яғни оның белгілерін (мысалы, шаштың түсін, қанның тобын, өсуді және т.с.с.) анықтайды. Гендер өзінде болатын ферменттер құрылымы және басқа жасушалық ақуыздар туралы ақпарат есебінен жасушалық метаболизмге бақылау жасайды. Ал ферменттер тірі организмдерде жүретін барлық химиялық реакцияларды басқаратын биокатализатор рөлін атқарады.Геннің құрылымы мен қызметін, Ген мен ферменттер арасындағы өзара байланысты әрі қарай тереңдете зерттеудің нәтижесінде “бір ген — бір полипептид” деген ұғым тұжырымдалды.Геннің қызметі туралы қазіргі көзқарастың қалыптасуына Америка ғалымдары Д. Бидл, Э. Тейтем (Татум) және С. Бензер жүргізген зерттеулердің әсері көп болды (1940 — 60).
№50 емтихан билеті
1. Құрылымды гендердің транскрипциясын бақылайтын элементтер.
Бір геннің транскрипциясы тұрақты жүрмейді, уақытқа байланысты жүреді, яғни гендер қосылып және ажырап отырады немесе гендер белсенді немесе белсенсіз халде болады.
Прокариоттарда гендерді қосатын өнімдерді индуктор, ал ажырататын өнімдер-репрессор деп аталынады.
Индуктор-бұл спецификалық реттелуші сигнал немесе бір реакцияның соңғы өнімі.
Репрессор – бұл белок. Ген, экспрессия индуктордың әсеріне деген жауапты күшейтеді. Бұл процесс индуцибельді ген деп аталады. Эукариот гендерінде индуктордың орнына реттеуші термині қолданылады. Реттеуші болып: белок, белсенді оттегі, металл саналады.
Геннің экспрессиялы реттелуі – бұл ДНҚ-ның әртүрлі бөлігіне немесе нүкте аймағына (сайттарға) белгілі өнімдер, мысалы белоктың спецификалы өзіндік қосылуын, транскрипцияның басталуы деп атаймыз. Сонымен гендердің экспрессиялы реттелуі дегеніміз қоршаған орта өзгерістеріне организмнің бейімделуі.
Реттеуші аймақ 2- элементтерден тұрады: 1-ші элемент базалы экспрессияның реттелу деңгейін қамтыйды /промотор/. 2-ші –қосымша экспрессиялы реттелу деңгейі.
2. Нуклеин қышқылдарының құрылымы мен қызметі.
Нуклеин қышқылдары биологиялық полимерлер класына жатады.Сондықтан ДНК және РНК –ның біріншілік құрылымы түсінігіне олардың мономерлік қалдықтарының құрылысы,мономераралық ковалентті байланыстардың химиялық табиғаты,полимер тізбектегі мономер звенолардың кезектесіп орналу тәртібі жатады. Нуклеин қышқылдарының екінші реттік құрылымы нуклеотидтік тізбектердегі көрші мономер буындар арасындағы әрекеттесу арқылы түзіледі, ал екі жақты спираль молекулалар немесе молекулалар учаскелері болған жағдайда екі жақты спиральда бір біріне қарама қарсы тұрған нуклеотид қалдықтары арасындағы әрекеттесулер арқылы түзіледі. Нуклеин олардың екінші, реттік құрылымдарының әртүрлі элементтеріне жататын нуклеотид қалдықтарының арасындағы әрекеттесулер есебінен ұйымдастырылады.қышқылдарының үшінші реттік құрылымы. Нуклейн қышқылдарның төртінші реттік құрлымы мен РНҚ молекуласына тән.РНҚ 2 бірдей суббірліктен тұрады.олардың байланысы Азотты негіздердей арасындағы комплематарлы әсерлесуі арқасында жузеге асады. Нуклейн қышқылы клеткаларыда қызметыне қарай шоғырланады.ДНҚ мен клетка ядросында,ал РНҚ ядро мен цитоплазмада көбінде еріген клеткада кездеседі. Нуклеин қышқылдары биологиялық тұрғыдан маңызды рөл атқарады. Олар тірі организмдердегі генетикалық ақпаратты сақтайтын және тасымалдайтын жасушаның (жасушаның) маңызды кұрам бөліктері болып табылады. Нуклеин қышқылдары ақуыз биосинтезіне қатысады және тірі организмдерде тұқым қуалаушылықты сақтап, оның бір ұрпақтан екінші ұрпаққа берілуін қамтамасыз етеді. ДНҚ жасуша ядросының хромосомасында (99%), рибосомаларда және хлоропластарда, ал РНҚ ядрошықтарда
3. Прокариоттар транскрипциясының ерекшеліктері.
Транскрипция –генетикалық ақпараттың ДНҚ молекуласын РНҚ молекуласына берілу процессі. Кейбір жағдайда барлық мРНҚ-ның молекуласы бір тізбектен көшіріледі. Екі тізбекте де РНҚ транскрипциясы екінші тізбек негізінде жүреді.Мұндай транскрипция әдісінің мәні реттеуші б.т. Прокариоттар гендері жұмыс істеуінің немесе экспрессиясының механизмі бүгінгі күні жақсы зерттелген және Е. СоІі бактериясы мен кейбір оның фагтарын көпжылдық іргелі молекулалық-биологиялық зерттеулер арқасында түсінікті бола бастады. Алдымен, прокариоттық геннің экспрессиясы деп осы генде иРНҚ синтезделуін (транскрипция кезеңі) және иРНҚ-ның рибосомадағы трансляциясын (белок синтезін) түсінетінін еске түсірейік.Белок синтезінің бірінші кезеңі — трапскрипцияның инициация процесінде РНҚ-полимераза ферменті промотор деп аталатын ДНҚ-ның ерекше бөлігін тануы керек, тек сонда ғана ДНҚ-ның қос тізбегі ажырап, иРНҚ синтсзделуі бастала алады. Демек, адам немесе жануар генінің бактерия клеткасында жұмыс істеп, қажет биотехнологиялық өнім (белок) түзуі үшін, ең алдымен, оның алдында бактериялық РНҚ-полимераза байланыса алатын прокариоттық күшті промоторболуы керек.
Транскрипцияның жоғары деңгейі плазмидалық репликацияның тұрақсыздығына әкелетін атап өту керек. РНҚ-ның ғана емес, әсіресе көптеген белоктардың мөлшерден тыс синтезделуі клетканың жойылуына әкелуі мүмкін. Осындай жағдай болмас үшін пәрменді транскрипцияланатын геннің соңында трапскрипцияны тиімді терминациялайтын (тежейтін) нүктелер болуы керек. Практикалық мақсат үшін реттелетін транскрипцияны қолдану ыңғайлы.
№51 емтихан билеті
1. Жалпы геном туралы түсінік.
Геном (ағылшынша genome, грекше genos — шығу, тек) — хромосомалардың гаплоидты (сыңар) жиынтығында шоғырланған гендердің бірлестігі. Геном терминін 1920 жылы неміс биологы Г. Винклер енгізді. Гаплоидты жиынтық көбінесе жынысжасушаларына тән, ал сомалық (дене) жасушаларында хромосомалардың диплоидты (екі еселенген) жиынтығы болады. Кейде хромосомалардың саны қалыпты диплоидты жағдайдан артып кетеді. Егер гаплоидты жиынтықтан Геном үш не төрт есе артық болса, триплоидты және тетраплоидты, ал бір Геном ағзада бірнеше рет қайталанса, автополиплоидты, ал әр түрлі біріккен ағза аллополиплоидты деп аталады. Хромосомалардың жиынтығы еселеніп, артқан сайын Геном саны да өсіп отырады. Әдетте диплоидты клеткада хромосомалар жұп болып келеді. Себебі, ұрықтану кезінде оның бір сыңары аналық гаметадан, екіншісі — аталық гаметадан беріледі, яғни бұл Геномдар сәйкес (гомологты) болады. Сөйтіп екі гаплоидты жасушадан бір диплоидты жасуша түзіліп, жаңа ағза қалыптасады. Әр хромосомада тізбектеліп орналасқан гендердің өзара дәл келуін екі Геномның сәйкестігі деп атайды. Туыстығы қашық буындарда барлық немесе бірнеше Геномдар арасында сәйкестік болмайды. Бұл тұрақтылық бұзылып, белгілі бір факторлардың әсерінен хромосома санының өзгеруін (мысалы, артып, не кеміп кетсе) геномдық мутация деп атайды.
Тірі ағзаларда хромосомалардың саны тұрақты болады. Мысалы адамда — 46, маймылда — 48, қиярда — 14, жүгеріде — 20, қатты бидайда — 28, жұмсақ бидайда — 42, дрозофила шыбындарында — 8, т.б. Организм эволюциялық дамуында неғұрлым жоғары сатыда тұрса, соғұрлым олардың Г-ында ДНҚ көбірек болады.
2. Нуклеин қышқылдарының зерттелу тарихы.
ДНҚ-ны 1868 жылы швейцар физиологы, гистологы және биологы Иоган Фридрих Мишер атты ғалым ашқан. Іріңдеген жасушалар қалдықтарынан ғалым құрамына азот пен фосфор кіретін бейтаныс затты тауып алады. Алғашында бұл жаңа зат нуклеин деген атқа ие болады. Кейіннен Мишер бұл заттың қышқылдық қасиет көрсететінің байқайды. Осыдан кейін бұл жаңа затты нуклеин қышқылы деп атайтын болған. Алғашында бұл бейтаныс заттың биологиялық қызметі белгісіз болды, көп уақытқа дейін ДНҚ ағзадағы фосфордың қоймасы болып есептелінді. Оған қоса, XX ғасырдың басында ғалымдар ДНҚ-ның ақпаратты тасымалдай алмайтындығын айтқан, себебі олар ДНҚ-ның ақпаратты тасымалдау үшін құрылысы біртүрлі деп есептеді.
Уақыт өте келе генетикалық ақпаратты наруыздар емес дәл осы ДНҚ тасымалдайтындығын дәлелдеді. Бұл ашылудың ең бірінші дәлелі О. Эвери, Колина Мак-Леода и Маклин Мак-Картидің бактериялардың трансформациясы(1944 жыл) тәжірбиесі болды.
Тіпті XX ғасырдың 50 жылдарына дейін ДНҚ-ның нақты құрылысы мен ақпаратты ұрпаққа белілуінің әдісі белгісіз болды.
ДНҚ-ның қос спиральінің құрылымын 1953 жылы Френсис Крик пен Джеймс Уотсон ұсынды. Олар модельді Морис Уилкисон мен Розалинд Франклиннің рентгенқұрылымды деректеріне және Чаргаффа ережелеріне сүйене отырып құрап шығарған.
3. Гендер экспрессиясын реттеудің практикалық маңызы.
№52 емтихан билеті
1.Эукариоттардағы трансляция процесінің реттелу механизмдері.
Трансляция — полипептид тізбегінің гендегі иРНҚ негі- зінде ақпаратқа сай түзілуі. Трансляция болашақ белокқа тән иРНҚ-на жазылған нуклеотидтер кезегін түзілетін белоктардың амин қьішқылдарының кезегіне ай- налдырады. Бұл жұмысқа иРНҚ-нан басқа рибосомалар, тРНҚ, аминоацил синтетазалар, белоктан тұратын инициация, элонгация және терминацияфакторлары қосылған күрделі құрамдар қатынасады
2. Гендер активтілігінің реттелуі.
Ген (грек. genos — тұқым, тек) — тұқым қуалаудың қандай да бір элементар белгісін қалыптастыруға жауапты материалдық бірлік. Генде жасушаның құрылымы мен қызметін анықтайтын генетикалық ақпарат болады. Бір организмнің Гендер жиынтығы оның генотипін құрайды.Ген терминін алғаш рет 1909 жылы Дания ғалымы В.Йогансен енгізді. Барлық Гендер ДНҚ-дан тұрады және әрбір жеке жасушадағы мыңдаған осындай Гендер жеке ДНҚ молекуларының үзіндісі түрінде емес, хромосома деп аталатын, ірі құрылымдық бірлік құрамында болады. Жасушаның бөлінуі кезінде бұл хромосомалар екі еселенеді және жаңа түзілген жас жасушаалар осындай ата-аналық Гендер жиынтығының көшірмесін алады. Соның нәтижесінде жасушааның барлық белгілері (қасиеттері) ұрпақтан ұрпаққа беріледі, яғни тұқым қуалайды. Әртүрлі органимздердегі Геннің орташа ұзындығы 1000 нуклеотид негіздерінің жұбынан құралады деп есептеуге болады. Мыс., жануарларда кездесетін SV-40 вирусындағы ДНҚ-ның ұзындығы 5000 нуклеотид, яғни ол 5 геннен; Т4 бактериофагы — 200, ішек бактериясы — 4600, ал адамның гаплоидты жасушасы 100000 — 500000 Гендерден тұрады. 1865 жылы чех ғалымы Г. Мендел организм белгілерінің жеке тұқым қуалайтынын және шағылысу (будандастыру) кезінде ұрпақтарында жоғалмай сақталатынын анықтады.
3. Тұқым қуалау ақпаратының жүзеге асырылуы.
Генетикалық ақпарат —организмдердің ұрпаққа беретін қасиеттері жөніндегі ақпарат.Генетикалық ақпарат нуклеин қышқылында оның негіздерінің кезегі түрінде жазылған. Ой жүзінде бұл әдіспен белокмолекуласының шексіз көп түрінің кодын жазуға болады. Генетикалық ақпарат бір ұрпақтан екінші ұрпаққа нуклеин қышқылының транскрипциясы арқылы беріледі. Генетикалық ақпарат өзгерісті, не өзгеріссіз түзетіліп сақталуы мүмкін. Бұған репарация, рестрикция, рекомбинация т. б. қатынасады.[1]
№53 емтихан билеті
1.ДНҚ репликациясының реттелу механизмдері.
Репликация ДНҚ молекуласының 2 еселеніп өсуі.Тірі организмде қос қабат спиральдың бөлінуі әрә түрлі ферменттер және белоктар арқылы іске асырылады.Бұл жағдайда,репликация жүру үшін біршама уақытта бөлінген тізбектербір-бірінен қашық тұруы қажет.Бұл қызметті және 2 бөлу процесін хеликаза(геликаза) атты АТФ тәуелді ферменттеркомплексі орындайды.Хеликаза тікелей репликация айырдың алдыңғы ДНҚмолекуласының қысқа учаскілеріне бөледі.Репликациялық айыр –аналық молекуласының бір мезгілде тарқату,синтез процесі жүретін учаскісі.ДНҚ мол-ң 2 тізбегінде 5'→3'бағытта синтезделеді.Жетекші тізбек үздіксіз 2 еселеніп өседі.ал артта қалған тізбек қысқа үзінділерден (Оказаки фрагмент) синтезделінеді,оның бағыты репликациялық айырдың жүру бағытына қарама-қарсы келеді.Хеликаза әсерінен қос тізбекті спиральді тарқату үшін энергия АТФ АДФ –КЕ фосфатқа гидролизденгенде бөлініп шығады.Негіздердің әрбір жұбын бөлу үшін шамамен 2АТФ-тың энергиясы жұмсалады.Бөлініп шыққан аналық ДНҚ-ң әрбір тізбегі белоктың бірнеше молекулаларымен тығыз байланысты.ОЛ белок ДНҚ байланыстырушы белок д.а.Бұл белок комплементарлы жұптардың түзілуіне және тізбекткердің қайта қосылуына бөгет болады. Соның себебімен ДНҚ-ның бөлінген әрбір тізбегі матрица ретінде репликацияға пайдаланалы.
2. Эукариоттық ДНҚ-полимеразаларға сипаттама.
А.Корнберг әріптестерімен бірге бірнеше рет ДНҚ биосинтезіне қатысушы ферментті ДНҚ-полимеразаны Е.соli клеткаларынан бөліп алады.(1955)Одан кейін ДНҚ полимераза II,ДНҚ-полимера III осы бактерия клеткасынан алынып сипаттама берілді.Эукариоттар клеткасында кем дегенде 3 ДНҚ-полимеразасы бар олар α-,β-,γ- деп аталған.α- ДНҚ-полимераза хромосома репликациясында түзілуінде негізгі фермент болып саналады.β-ДНҚ- полимераза ДНҚ-полимеразаI сияқты,ДНҚ молекуласының бұзылған учаскілерін жөндеуге қатысады.α-және β- ДНҚ полимераза ядрода,ал γ- ДНҚ-полимеразасы митохондрияда болады.Эукариот клеткаларында барлық ДНҚполимераза олигомерлы белоктар.
3. Рибосомалық РНҚ-ның қызметі.
р-РНҚ басқа РНҚ қарағанда мөлшер жағынан басым болады.80 пайыз шамасындай.Молекулалық массасы да өте жоғары болады.р-РНҚ рибосома құрылымын түзейтін компоненттің біреуі. Эукариот клеткаларында 4түрлі р-РНҚ,прокариоттарда 3 түрлі болады.РНҚ полимераза I-дің әсерінен ұзын транскриптер түзіледі,процессинг процесінен кейін «піскен» рРНҚ құралады.Эукариоттар рибосомасына құрамына кіретін 4 түрлі рРНҚ молекулаларының ұзындық,шөгу коэффиценті және нуклеотидтер санынан бір-бірінен айтарлықтай өзгешеліктері бар.рРНҚ молекулалары күрделі 2-шілік,3- шілік құрылымдар түзеді.Комплементарлы учаскілерінде ілгектер түйреуіштер құрап,рибосомаға ерекше пішін береді.
№54 емтихан билеті
1. Белок биосинтезінің реттелу механизмі.
Белок биосинтезінің жылдамдығын бір жағынан генетикалық аппарат басқарса 2-ші жағынан сыртқы орта факторлары басқарады.Эукариот клеткаларында белок биос. Реттелуінің 2 механизмі болуы мүмкін.Олар: транскрипция деңгейінде реттелу және трансляция деңгейінде реттелу. Транскрипция деңгейінде реттелу. ДНҚ хромосома ішкі жағында, ал белоктар сыртқы тыс жағында «орама» түрінде орналасқан.Гендердің активтенуі ДНҚ молекуласының учаскесінің белоктан босап шыгуына байланысты осы жағдай мРНҚ транскрипциясына алып келеді.Барлық гормондар дерлік транскрипция жылдамдығын арттырады яғни мРНҚмөлшерін арттырады сөйтіп белок биосинтезін ұлғайтады. Трансляция деңгейінде реттелу. Трансляция негізгі 3 сатыдан тұрады.Олар: инициация,элангация, терминация.Белок син. Реттелуі осы сатыларының әрқайсысының кезінде-ақ орналасады.Мұнда клеткалардың амин қышқылдарымен қамтамасыз етілуі маңызы зор.Белгілі бір амин қышқылы жетіспеген кезде оған сәйкес аминоацил- тРНҚ түзілуі тоқталады. Оның салдарынан трансляция тежеледі.
2. Рибосомалар,оның құрылысы және қызметі.
Рибосомалар- клетканың белок синтездеуші молекулалық апараты болып есептеледі,мұнда ДНҚ-дан м-РНҚ алып келген жоспар бойынша белок түзіледі.Химиялық табиғаты жағынан қарағанда:рибосома- рибонуклейн қышқылдарынан және белоктардан құралған рибонуклеинпротеиндық комплекс.Рибосоманың құрылымы өте күрделі, бірнеше р-РНҚ- дан және көптеген әртүрлі белоктардан құралған.Рибосома бір-біріне тең емес 2 бөліктен құралған.Эукариоттардың тұтас рибосомасы 80S деп,үлкен суббөлігі 60S,кіші 40S. Прокариот тұтас риб.70S, үлкен 50S,кіші 30S(S- Сведберг өлшемі) Олар жасуша цитоплазмасындағы Mg иондарының концентрациясына байланысты диссоциацияланып (молек. ыдырауы) және реассоциацияланып (молекулалардың қайта бірігуі) отырады. Рибосомалар жасушада ақуыз синтезіне қатысатын құраушыларды ұстап тұру, оларды бір-бірімен байланыстыру қызметін атқарады.. Рибосомадағы ферменттер, магний мен кальцийдің тұздары зат айналым өзгерістеріне қатысады. Рибосомада минералдық элементтерден азот,фосфор, темір және мыс кездеседі.
3. Генетикалық кодтың қасиеттері.
Генетикалық код — тірі организмдерге тән нуклеин қышқылдары молекуласындағы тұқым қуалаушы (генетикалық) ақпараттың нуклеотидтер тізбегі түріндегі біртұтас “жазылу” жүйесі. Бұл — барлық тірі организмдерге ортақ заңдылық. Генетикалық код бірлігі — ДНҚ мен РНҚ молекуласындағы 3 нуклеотид (триплет) тізбектерінен тұратын кодон (аРНҚ нуклеотидтерінің триплеттері) болып табылады. Гендегі кодондар тізбегі осы генді “жазатын” (кодтайтын) ақуыздағы амин қышқылдар тізбегін анықтайды. Клеткадағы генетикалық код екі сатыда іске асады:1.транскрипция сатысы ядрода жүреді және ДНҚ-ның сәйкес бөліктерінде ақпараттық (информациялық) рибонуклеин қышқылдарының молекулалары (аРНҚ) жасалады. Сонымен қатар, ДНҚ нуклеотидтер тізбегі аРНҚ нуклеотидтер тізбегі ретінде қайта жазылады; 2.трансляция сатысы цитоплазмада, ақуыз синтезделетін рибосомада жүреді. Сондай-ақ, аРНҚ нуклеотидтер тізбегі, полипептидтер құрайтын амин қышқылдар қалдықтарының белгілі бір тізбегіне көшеді. Генетикалық кодтың бір ерекшелігі, әмбебап екендігі, яғни барлық организмдерде белгілі бір 3 нуклеотид (триплет) белгілі бір амин қышқылдарын “жазады” (кодтайды). Бір амин қышқылы бірнеше триплетпен “жазылуы” (кодталуы) мүмкін. Кодондар арасында “үтір” болмайды, яғни олар бір-бірінен бөлінбеген. Ол бір геннің аймағында белгіленген нүктеден бастап, бір бағытта есептелінеді. 64 кодонның 61-і ақуыз құрайтын 20 амин қышқылдарын “жазады” (кодтайды), ал қалған үш “нонсенс” (мағынасыз) кодондар (УАГ, УАА және УГА) полипептид синтезін аяқтайтын “нүкте” қызметін атқарады. Олар ақуыз биосинтезінінің аяқталғанын білдіреді.
№55 емтихан билеті
1.Гендік инженерия, жалпы түсінік.
Гендік инженерия, генетикалық инженерия [1] — генетикалық және биохимиялық әдістердің көмегімен түраралық кедергілері жоқ, тұқым қуалайтын қасиеттері өзгеше, табиғатта кездеспейтін жаңа гендер алу; молек. биологияның бір саласы. Гендік инженерия әр түрлі организмдер геномының бөлігінен рекомбинатты ДНҚ құрастырумен қатар, ол рекомбинатты молекулаларды басқа ағза геномына енгізіп, жұмыс істеуін (экспрессиясын) қамтамасыз етеді. Гендік инженериядағы тұңғыш тәжірибені 1972 ж. американ биохимигі Т. Берг (Нобель сыйл. лауреаты) іске асырды. Ол маймылдың онноген вирусы SV-40-тың толық геномын, бактериофаг — L геномының бір бөлігін және Е. Colі бактериясының галактоза генін біріктіру арқылы рекомбинантты (гибридті) ДНҚ алды. 1973 — 74 ж. Америка биохимиктері С. Коэн, Г. Бойер, т.б. түрлі ағзалардан бөліп алынған генді бактерия плазмидасының құрамына енгізді. Бұл тәжірибе басқа организмдер гендерінің жаңа ағза ішінде жұмыс істей алатынын дәлелдеді. Жануарлар клеткаларымен жүргізілген тәжірибелерде бір клетканың ядросын екіншісімен алмастыруға, екі немесе бірнеше эмбриондарды қосып біріктіруге, оларды бірнеше бөлікке бөлшектеуге болатыны анықталды. Мыс., генотиптері әр түрлі тіндердің клеткаларын біріктіру арқылы тышқанның аллофенді особьтары (фенотипі әр түрлі дарабастар) алынды. Гендік инженерия-ның теориялық негізіне генетикалық кодтың әмбебаптылығы жатады. Бір ғана кодтың (триплиттің) әр түрлі ағзадағы белок молекулаларының құрамына енетін амин қышқылдарын бақылай алатындығына байланысты, ДНҚ молекуласының кез келген бөлігін басқа бөтен клеткаға апарып салу, яғни молек. деңгейде будандастырылу теориялық тұрғыдан алғанда мүмкін екені анықталды. Жануарлар, өсімдіктер және микроорганизмдер гендерінің қызметін қолдан басқаруға болатындығы дәлелденді. Ауыл шаруашылығында өсімдіктің атмосфералық азотты өзіне жинақтап алуы — үлкен мәселе. Осыған байланысты 1970 жылдары азотты фиксациялауға қабілеті жоқ пішен таяқшасына азотты жинақтай алатын, басқа бір бактерияның гені салынып, азотты жинақтау қасиетіне ие болды. Мед. саласында жаңа гендерді енгізу арқылы тұқым қуалайтын ауруларды емдеуге болады. Қазіргі кезде ауру адамдардан зат алмасудың 1000-нан аса әр түрлі тұқым қуалайтын өзгерістері табылған.
2. Мутациялық өзгергіштіктің түрлері.
Мутациялық өзгергіштік – дегеніміз организмнің генотипінің өзгеруіне байланысты болатын, яғни тұқым қуалайтын өзгергіштік. Мутациялар әр түрлі бағытта жүзеге асады. Олардың көпшілігі организмнің тіршілік қабілетін кемітіп жібереді. Кейде өлімге душар етеді, оны летальді мутация деп атайды. Жалпы мутацияны эволюциялық процесстің алғашқы материалдық негізі деп есептеу керек. Бірақ жеке мутациялар түрді өзгерте алмайды. Мутация жыныс жасушаларындағы және хромосома аппаратындағы өзгерістеріне байланысты бірнеше типке бөлінеді. Мутацияның типтері. Мутациялық процестің өзін секірмелі түрде, яғни кенеттен пайда болатын және индукциялық деп бөледі.. Мутация сыртқы орта факторларының әсерінен пайда болады, оларды мутагендер деп атайды. Мутагендердің үш түрі кездеседі. Олар: физикалық, химиялық және биологиялық мутагендер. Физикалық мутагендерге радиоактивті сәулелер, ультракүлгін сәулелер, лазер сәулелері және т.б. жатады. Химиялық мутагендерге колхицин, этиленимин, никотин қышқылы және т.б. химиялық қосылыстар жатады. Гендік мутация. Мутацияның мұндай түрі жекелеген гендерде болады және жиі кездеседі. Гендік мутация ДНҚ молекуласындағы нуклеотидтердің орналасу ретінің өзгеруіне байланысты болады. Мысалы, ДНҚ құрамындағы қатар тұрған екі нуклеотидтің орын алмастыруы немесе бір нуклеотидтің түсіп қалуы мүмкін. Соның салдарынан генетикалық код өзгереді де, тиісті белок синтезделмей қалады немесе синтезделген белоктың қасиеті өзгереді. Ол ақыр аяғында келіп, организм белгісінің өзгеруіне апарып соғады. Гендік мутацияның нәтижесінде жаңа аллельдер пайда болады. Оның эволюция мен селекция үшін үлкен маңызы бар. Мысалы, селекцияда өсімдіктердің жаңа сорттарын, жануарлардың тұқымдарын және микроорганизмдердің жаңа түрлерін алу үшін қажетті материал ретінде пайдаланылады. Гендік мутацияның негізінен екі түрін ажыратады: Азотты негіздердің орын ауыстыруына және бір азотты негіздердің ДНҚ құрамынан түсіп қалуына немесе үстеме келіп қосылуына байланысты болатын мутациялар. Мұндай өзгерстердің нүктелік мутация деп те атайды. Геномдық мутация – дегеніміз жасушадағы хромосомалар санының өзгеруіне байланысты организмнің белгілері мен қасиеттерінде пайда болатын өзгергіштікті айтады. Геном дегеніміз гапплидті хромососадағы гендердің жиынтығы.Енді осы геномдық мутациянын пайда болу жолын қарастырайық. Хромосома санының тұрақтылығын және оның ұрпақтан ұрпаққа берілуін қамтамасыз ететін жасушаның бөлуін механизмдері митоз мен мейоз екендігі белгілі. Бірақ кейбір жағдайда бұл механизмдер бұзылады да, хромосомалар жасушадағы екі полюске теңдей ажырамайды. Соның салдарынан хромосома саны өзгерген жасушалар пайда болады. Геномдық.мутация тұтас гаплоидты жиынтықтың немесе хромосомалар санының көбеюіне немесе азаюына байланысты.
. Гендер активтілігінің гормональдық реттелуі.
Ген экспрессиясы (лат. expressio - əйгілену, ағ. gene expression) – ДНҚ
молекуласындағы нуклеотидтер тізбегіндегі генетикалық информацияны жүзеге
асыру. Ол: транскрипция, процессинг, трансляция сияқты негізгі үш сатыдан
тұрады.
№56 емтихан билеті
1. Гендік мутацияға сипаттама
Гендік мутация деп ДНҚ молекуласының белгілі бір бөлігінде нуклеотидтердің қатар тізбегінің өзгеруін айтады. Ол молекулалық деңгейде өтеді, микроскоп арқылы көрінбейді. Мутация нәтижесінде ағза биохимиялық, физиологиялық, морфологиялық өзгерістерге ұшырайды. Организмдегі бұл өзгерістер бірден немесе біраз уақыттан кейін біртіндеп байқала бастайды. Полиплоидты мутанттардың клеткалары мен органдарының көлемі ұлғайып, хромосома жиынтығы жұп болса, оның ұрпақ беру қабілеті сақталады, ал тақ болса бұл қабілеті сақталмайды. Гендік мутация кезінде ағза үлкен өзгеріске ұшырайды. Кейде бір геннің өзгеруінен ағзаның бірнеше белгі-қасиеттері өзгереді (плейотропия). Гендік мутация доминантты (басыңқы), жартылай доминантты және рецессивті (басылыңқы) болады. Хромосомалық және гендік мутациялардың себебі көпке дейін белгісіз болып келді. Бұл өзгерістер ағзаға әр түрлі физикалық, химиялық факторлар – мутагендердің әсер етуінен пайда болады. Мысалы, радий сәулелерінің саңырауқұлақта тұқым қуалайтын өзгеріс тудыратындығын 1925 ж. орыс ғалымдары Г.А. Надсон (1867 – 1940) мен Г.С. Филлипов ашты. 1927 ж. АҚШ ғалымы Г.Меллер (1890 – 1967) жасанды мутацияның рентген сәулелерінің әсерінен болатынын тәжірибе жүзінде дәлелдеді. АҚШ генетигі С.Райт (1889 – 1988), орыс ғалымы С.С. Четвериков (1880 – 1959), ағылшын биологі Дж. Холдейн (1892 – 1964) қазіргі популяциялық генетиканың негізін салып, мутацияның эволюциялық мәнін ашты. Мутация көпшілік жағдайда ағза үшін зиянды болып келеді. Түрлі тұқым қуалайтын аурулар мен кемістіктерді тудырып, кейде тіпті өлімге душар етеді. Сонымен қатар кейбір мутациялар ағзаға пайдалы өзгерістер де алып келеді. Мысалы, гендік мутация (табиғи және қолдан сұрыптауға қажетті негізгі материал береді) өсімдіктер, жануарлар және микроазғаларды сұрыптау жолымен жаңа түрін алғанда кейбір қасиеттерін жақсартады.
2. Эукариот геномының экзон-интрондық құрылымы.
Эукариот геномынын қызметінде колинеарлы емес принцип сайкес келеді. Эук-ң гені ДНҚ тизбегинин 2 тізбегінен құралғаны мәлім болды.Американ генетигі У.Гильберттің ұсынысы бойынша олар экзон жане интрон деген атауларга ие болды.Экзон д\з-ақуыздардың аминқышқылдары туралы ДНК-нын акпараттык бөлігі. Будан эукариот геномынын үзілмелі екенін көреміз.әдеттегі гендер (экзон)интрондармен бөлігген.Мұндай үзілмеліліктің өзі маңызды болуы мүмкін:мысалы:овальбумин генінде 8экзон және 7интрон бар,гемоглобин тізбегі генінің 3 экзоны 2интронмен бөлінген, ал иммуноглобулиннің ауыр тізбегінің генінде 4 интронды және 5-тен көп экзонды ажыратады.Бұдан басқа интрондардың басым көпшілігі экзондардан ұзын болып шықты:кәдімгі гендереде экзондардың жалпы ұзындығы 1000н.ж болса, ал интрондардын орт.ұзынд. 5000-20000н.ж аралығында өзгереді. Геннің интрон бөлігінің барлық ‘оқылу’ шекараларында терминация кодондары бар,сондықтан далипептидтік тізбекті коделей алмайды.Эукариот клеткасында тіршілік ететін кейбір вирустардың гендері де экзон және интрон бөліктерден құралған.Жалпы вирус генінің ұйымдастырылуы өзі паразиттік тіршілік ететін жануарлардын гендерімен ұқсас.Сонымен эукариоттардын көптеген гендерінде интрондар бар6алайда олардын бірқатар гендерінде (рибосомада,гистонда) интрондар жоқ.1978ж эукариоттардын үзілмелі генінен өтетін ақуыз синтезін бактериялармен салыстырғанда косымша кезенды керек ететіні белгілі болды. Ең алдымен РНҚ-полимераза II ферменті өз функцияларын атқаруға дайын емес РНҚ тізбегін синтездейді және ол ДНҚ-ға толық сәйкес синтезделеді. Демек мұндай иРНҚ әртүрлі тізбектерден экзон және интрондардан құралған,соған байлансты гетерогенді ядролық РНҚ д.а. гяРНҚ-дан интрон бөліктері үзілгеннен кейін тек экзондардан куралган және цитоплазмада трансляция процесінде өз қызметін атқара алатын жетілген иРНҚ пайда болады.Бірнеше ферменттік жүйелер арқылы гяРНҚ-ның жетілген иРНҚ-ға айналуы процессинг деп,ал ондағы интрон бөліктерінің арнайы ферменттік жүйелер арқылы үзілуі сплайсинг д.а.М.Эдмондс және Дж.Браверман 1971ж эук-ң барлық иРНҚ-сы мен гяРНҚ-сының шамамен жартысының 3`ұштарында 50-ден 200-ге тек аденин нуклеотидтерінен ғана құралған кесінділерді тапты. Бұл соңғы бөліктер поли(А)-ұш деп белгіленеді.Мұндай поли (А) кесінділер геномда (ДНҚ-да) табылмады. Бұдан олар гяРНҚ-ға,кейін иРНқ-ға транскрипциядан соң жалғанатыны аян болды. Полиаденилдеу сплайсинг үшін сигнал болып саналады.
3. Хромосома және геном деңгейіндегі генетикалық инженерия.
Генетикалық инженерия — генетикалық және биохимиялық әдістердің көмегімен түраралық кедергілері жоқ, тұқым қуалайтын қасиеттері өзгеше, табиғатта кездеспейтін жаңа гендер алу; молек. биологияның бір саласы. Гендік инженерия әр түрлі организмдер геномының бөлігінен рекомбинатты ДНҚ құрастырумен қатар, ол рекомбинатты молекулаларды басқа организм геномына енгізіп, жұмыс істеуін (экспрессиясын) қамтамасыз етеді. Гендік инженериядағы тұңғыш тәжірибені 1972 ж. американ биохимигі Т. Берг (Нобель сыйл. лауреаты) іске асырды. Ол маймылдың онноген вирусы SV-40-тың толық геномын, бактериофаг — L геномының бір бөлігін және Е. Colі бактериясының галактоза генін біріктіру арқылы рекомбинантты (гибридті) ДНҚ алды. 1973 — 74 ж. Америка биохимиктері С. Коэн, Г. Бойер, т.б. түрлі организмдерден бөліп алынған генді бактерия плазмидасының құрамына енгізді. Бұл тәжірибе басқа организмдер гендерінің жаңа организм ішінде жұмыс істей алатынын дәлелдеді. Жануарлар клеткаларымен жүргізілген тәжірибелерде бір клетканың ядросын екіншісімен алмастыруға, екі немесе бірнеше эмбриондарды қосып біріктіруге, оларды бірнеше бөлікке бөлшектеуге болатыны анықталды. гендік инженерия жаңа ДНҚ алуға мүмкіндік береді. Мысалы, ұштары жабысқақ ДНҚ фрагменттерін алуға болады. Ол үшін ажыратылған немесе жасанды синтезделген ДНҚ учаскелерін эндонуклеаза ферменттерімен өңдейді, ол оның ұштарын қысқартады. Сосын басқа ферменттің көмегімен — полинуклеотидтрансферазаның көмегімен әлгі учаскелердің ұштарына аденин және тимидин нуклеотидтер жалғанады. Алынған рекомбинатты ДНҚ молекуласын бөтен генді бактермальды клеткаға ауыстыру үшін қолданады. Мұндай схема инсулин, интерферон, иммуноглобулин және басқалардың гендерін алу үшін қолданылған. Айта кету керек, генді жасау және оны қожайын-ағзаның хромосомасына енгізу оның синтез өнімін алуға бірден мүмкіндік бермейді. Ген қызмет етуі үшін, олар мәлімет кодталған учаскемен бірлескен тағы реттегіш учаскеге ие болуы керек.
№57 емтихан билеті
1. РНҚ-ның түрлері және қызметтері.
Рибонуклеин қышқылдары рибосомалық (рРНҚ), ақпараттық (аРНҚ) және тасымалдаушы (тРНҚ) болып бөлінеді. Рибонуклеин қышқылы тізбегі бірнеше ондаған нуклеотидтерден бірнеше мыңдаған нуклеотидтерге дейін созылатын біржіпшелі полинуклеотидтерден тұрады. Организмде РНҚ ақуыздармен кешенді байланысқан рибонуклеотидтер түрінде болады. РНҚ генетикалық ақпараттың жүзеге асуы мен ақуыз синтезіне қатысып, барлық тірі организмдерде аса маңызды биологиялық рөл атқарады. Көптеген вирустарда РНҚ-н жалғыз нуклеинді компонент (құраушы) құрайды. Осындай РНҚ вирустарда РНҚ биосинтезімен қатар ДНҚ биосинтезінде де матрица рөлін атқара алады (кері транскриптаза). Бактериялар, өсімдіктер және жануарлар жасушаларында құрылымы, метаболизмі және биол. қызметтері әр түрлі РНҚ типтері кездеседі. Мысалы, рРНҚ рибосоманың құрамына еніп, жасушадағы РНҚ-ның негізгі массасын құрайды және көлемі, құрылымы түрлі организмдерде әр түрлі болады. Клеткада негізінен рРНҚ-да ақуыздың биосинтезі жүреді; тРНҚ жасушада амин қышқылдары қалдықтарын жалғастырып алып, оны ақуыз синтезі өтіп жатқан жерге тасымалдайды. Әрбір амин қышқылының өзіне сай арнайы тРНҚ (әдетте бірнеше) болады. Барлық тРНҚ жоңышқа жапырағына ұқсас макромолекулалы құрылымға ие. Олардың рибосомаға және аРНҚ-на жабысатын, үш нуклеотидтен тұратын (антикодон) және амин қышқылы қалдығын жалғастыратын аймақтары бар. РНҚ-ның барлық түрлері жасушада ДНІ матрицасында синтезделеді, соның нәтижесінде ДНҚ-ндағы дезоксирибонуклеотидтер тізбегінде комплементарлы рибонуклеотидтер тізбегі құрастырылады, мұны транскрипция процесі деп атайды. Клетка ядросында матриц. РНҚ-ның (мРНҚ) бастамасы болып келетін алып молекулалар табылған, олардың көп бөлігі ядрода ыдырайды да, аз бөлігі цитоплазмаға өтіп, нағыз мРНҚ-ын құрайды.
2. Гендердің жіктелуі.
Ген (грек. genos — тұқым, тек) — тұқым қуалаудың қандай да бір элементар белгісін қалыптастыруға жауапты материалдық бірлік. Генде жасушаның құрылымы мен қызметін анықтайтын генетикалық ақпарат болады. Бір организмнің Гендер жиынтығы оның генотипін құрайды.Ген терминін алғаш рет 1909 жылы Дания ғалымы В.Йогансен енгізді. Барлық Гендер ДНҚ-дан тұрады және әрбір жеке жасушадағы мыңдаған осындай Гендер жеке ДНҚ молекуларының үзіндісі түрінде емес, хромосома деп аталатын, ірі құрылымдық бірлік құрамында болады. Жасушаның бөлінуі кезінде бұл хромосомалар екі еселенеді және жаңа түзілген жас жасушаалар осындай ата-аналық Гендер жиынтығының көшірмесін алады. Соның нәтижесінде жасушааның барлық белгілері (қасиеттері) ұрпақтан ұрпаққа беріледі, яғни тұқым қуалайды. Әртүрлі органимздердегі Геннің орташа ұзындығы 1000 нуклеотид негіздерінің жұбынан құралады деп есептеуге болады. Мыс., жануарларда кездесетін SV-40 вирусындағы ДНҚ-ның ұзындығы 5000 нуклеотид, яғни ол 5 геннен; Т4 бактериофагы — 200, ішек бактериясы — 4600, ал адамның гаплоидты жасушасы 100000 — 500000 Гендерден тұрады. 1865 жылы чех ғалымы Г. Мендел организм белгілерінің жеке тұқым қуалайтынын және шағылысу (будандастыру) кезінде ұрпақтарында жоғалмай сақталатынын анықтады. Будандардың бірінші ұрпағында ата-ананың біреуінің ғана белгісінің басым болуы доминанттық деп аталады. Генетикада Гендерді латын әліпбиінің әріптерімен белгілеу қалыптасқан, мыс., доминантты Генді бас әріппен (А), ал рецессивті (басылыңқы) Генді кіші (а) әріппен белгілейді. Микроорганизмдерде белгілі бір қосылыстар синтезіне жауапты Гендерді сол қосылыстар атауының алғашқы әріптерімен және “+” (қосу) белгісімен белгілейді, мыс., hіs+ — гистидин Гені, leu+ — лейцин Гені, тағыда басқа Гаметалардың түзілуі мен ұрықтану процестеріндегі әртүрлі Гендер бойынша белгілердің тәуелсіз ажырауы мен гомологтық емес хромосомалар әрекетінің арасындағы қатарластық (параллелизм), тұқым қуалаушылықтың хромосомалық теориясының негізін қалады. Бұл теория бойынша Гендер хромосомаларда тізбектеле орналасады да, олар тұқым қуалаушылықтың материалдық негізін қалайды (қ. Мейоз). Жасушадағы ақуыздың синтезделуі және олардың қарым-қатынасы туралы ақпарат тек Гендерде болады, яғни әрбір Ген белгілі бір ақуыз(полипептидті тізбек) синтезіне жауапты. Ақуыз синтезін бақылай отырып, Ген организмдегі барлық химиялық реакцияларды басқарады, яғни оның белгілерін (мысалы, шаштың түсін, қанның тобын, өсуді және т.с.с.) анықтайды. Гендер өзінде болатын ферменттер құрылымы және басқа жасушалық ақуыздар туралы ақпарат есебінен жасушалық метаболизмге бақылау жасайды. Ал ферменттер тірі организмдерде жүретін барлық химиялық реакцияларды басқаратын биокатализатор рөлін атқарады.Геннің құрылымы мен қызметін, Ген мен ферменттер арасындағы өзара байланысты әрі қарай тереңдете зерттеудің нәтижесінде “бір ген — бір полипептид” деген ұғым тұжырымдалды.Геннің қызметі туралы қазіргі көзқарастың қалыптасуына Америка ғалымдары Д. Бидл, Э. Тейтем (Татум) және С. Бензер жүргізген зерттеулердің әсері көп болды (1940 — 60).
3. Прокариоттық ДНҚ-полемеразаларға сипаттама.
Прокариоттық ДНҚ полимеразалар Ішек таяқшасы бактериасында және басқа прокариоттарда қызмет атқаратын ДНҚ полимеразалардың үш типі белгілі. ДНҚ полимеразалар 1 және полимераза 2 ДНҚ репарациясына қатынасады, ал ДНҚ полимераза 3 ДНҚ репликациясына қатынасады. ДНҚП күрделі фермент, оның 5 активті сайттары бар: 1. матрицалық сайт 2. праймерлік сайт 3. 5!-3! Ыдыратушы сайт немесе экзонуклеазалық сайт 4. Нуклеотит- үшфосфат сайт 5. 5!-3! Эндонуклеазалық сайт. ДНҚП 1 ферментінің атқаратын қызтеті синтездік активтілік көрсетіп, нуклеотиттерді ретімен қосып ДНҚ тізбегін өсірумен қатар экзонуклеозалық активтілігіне байланысты тізбектердегі қателіктерді жөндеп, түзету болып табылады. ДНҚП 2 активтілігі жағынан ДНҚ 1ге ұқсас болсп келеді, бірақ нуклеотиттерді қосып, ұзартып өсіру үшін тізбектің 3! Бос ұшына ғана әсер ете алады. ДНҚП ДНҚ репликациясында маңызды роль атқарады. Бұл мультимерлік фермент немесе холофермент суббірліктен тұрады.
№58 емтихан билеті
1. Эукариоттық ДНҚ-полемеразаларға сипаттама.
Эукариоттық ДНҚ полимеразалар 1. ДНҚ полимераза альфа цитоплазмалық немесе үлкен полимераза деп те аталады, ол ядрода және цитолпазмада кездеседі. 2. ДНҚ полимераза бета нулеазалық полимераза кіші полимераза, тек омыртқылыларға тән фермент. 3. ДНҚ полимераза гамма митохондриялық полимераза, ядода синтезделеді. 4. ДНҚ полимераза дельта сүтқоректердің жасушаларынан табылған және PSNA белогіна тәуелді; ол ДНҚ синтезінің жылдамдығы мен қарқындылығын қамтамасыз етеді. 5. ДНҚ полимераза эпсилон қызметі бойынша дельта ДНҚ полимеразаға ұқсас, ашытқы саңырауқұлақтары мен сүтқоректілердің жасушаларында кездеседі ДНҚ молекуласындағы реаликация басталатын нүтеде А- Т жұптары көбірек, арнайы нуклеотиттер жүйесі орналасқан болады.
2. Теломеразалар әрекетінің механизмі.
Теломеразалар – жасушаның әр бөлінуінде қысқарған хромосома ұштарын қалпына келтіруші фермент.
Барлық жасушаларда ДНҚ-ның толық репликацияланбаған учаскелері қалпына келуі қажет. Бұл қызметті теломеразалар атқарады.
Теломеразалар әрбір теломерлердің G-тізбегін ұзартады. Теломеразалармен 450 нуклеотидтерден тұратын теломеразалық РНҚ байланысқан. Оның ортаңғы қысқа учаскесі 1,5 теломерлік қайталануға комплиментарлы болады. Осы РНҚ –ның сол жағындағы триплет (АУЦ) ДНҚ-ның G-тізбегінің шеткі теломерлік жартықайталанумен байланысу үшін пайдаланылады. Қалған гексонуклеотид (ЦЦААУЦ) G-тізбекті 3′ ұщынан ұзарту үшін матрица ретінде қызмет атқарады. Теломеразалардың қызметі: қысқа, жаңадан синтезделген тізбекті ұзартпай, ескі матрицалық ұзын тізбекті ұзартады.
3. Транскрипция процесінің ерекшеліктері.
ТРАНСКРИПЦИЯ (лат. transcrіptіo – қайта көшіріп жазу) – тірі жасушалардағы рибонуклеин қышқылының биосинтез процесі. Ол дезоксирибонуклеин қышқылы (ДНҚ) матрицасында жүреді. Транскрипция аденин,гуанин, тимин және цитозиннің қайталанбалы тізбегінен тұратын ДНҚ молекуласындағы генетикалық ақпараттың іске асуының бірінші кезеңі. Транскрипция арнайы ДНҚ және РНҚ полимераза ферменті арқылы жүреді. Транскрипция нәтижесінде РНҚ молекуласының полимерлі тізбегі түзіледі. Бұл тізбек ДНҚ молекуласының көшірілген бөлігіне комплементарлы болады.
Транскрипция бірлігін атқаратын қызметі бір-біріне байланысты ферменттер синтезін анықтайтын гендер тобын ``оперон`` деп атайды. Прокариоттарда ол функционалды байланысқан бірнеше геннен, ал эукариоттарда тек бір ғана геннен тұрады. РНҚ-полимераза ферменті оперонның бастапқы бөлігін (промотор) “таниды”, онымен байланысып, ДНҚ молекуласының қос тізбегін ширатады. Осы жерден бастап мономерлі нуклеотидтер комплементарлы шартқа (принципке) сай РНҚ молекуласын түзеді. РНҚ-полимераза ферментінің ДНҚ-матрицасымен жылжуына байланысты синтезделген РНҚ молекуласы алшақтай береді де, ДНҚ-ның қос тізбегі қайта қалпына келеді. РНҚ-полимераза көшірілетін бөліктің соңына жеткенде (терминатор) РНҚ молекуласы матрицадан ажырайды. ДНҚ молекуласының әр түрлі бөліктеріндегі көшірмелер саны жасушаның қандайда болмасын ақуызды қажетсінуіне және қоршаған орта жағдайларына байланысты болады. Транскрипция процесінің реттелуін зерттеу молекулалық биологияның маңызды міндеттерінің бірі болып саналады. Ақпараттың көшірілуі ДНҚ молекуласынан РНҚ-ға ғана емес, сондай-ақ, кері бағытта, РНҚ-дан ДНҚ-ға да көшірілуі мүмкін. Мұндай кері Транскрипция құрамында РНҚ молекуласы бар ісік тудыратын вирустарда болады. Олардың құрамында жасуша зақымданғаннан кейін вирустың РНҚ-сын ДНҚ тізбегін синтездеуге матрица ретінде қолданатын фермент болады. Соның нәтижесінде ДНҚ молекуласының бір тізбегі, яғни ДНҚ – РНҚ гибриді түзіледі. Алғашқы РНҚ молекуласының барлық ақпаратын алып жүретін вируспен зақымданған қос спиральды ДНҚ молекуласы жасушаның хромосомасына еніп, қатерлі ісік тудырады. Кері Транскрипцияның ашылуы Ресей ғалымы Л.А. Зильбер (1894 – 1966) ұсынған қатерлі ісік вирусты-генетикалық теорияның дұрыс екенін дәлелдеді. Кері Транскрипция қалыпты жасушаларда ақпараттың жинақталуында және оның іске асуында (мысалы, эмбрионды даму кезеңінде) маңызды рөл атқаруы мүмкін
№59 емтихан билеті
1. РНҚ-полемеразаның құрылымы.
Бактериялар клеткасында тек қана бір РНҚ-полимераза барлық РНҚ-дың түзілуін катализдейді. Эукариоттар клеткасында бұл қызметті үш түрлі РНҚ-полимеразалар – РНҚ-полимераза I,II және III атқарады.
Бактерия клеткасында РНҚ-полимеразадан басқа тағы бір фермент бар, ол ДНҚ репликациясы үшін керек, РНҚ-қордың синтезін катализдейді. Ол бір полипептидтік тізбектен құралған, М 60000.
E.coli РНҚ – полимеразасы голофермент (комплекс) болып саналады, М 390000, бір-бірімен молекулалық масса айырмасы бар 6 суббөлшектерден тұрады.
| Голоферментінің суббөліктері | Молекулалық массасы |
| α (екеу) | |
| β (біреу) | |
| β’ (біреу) | |
| ω (біреу) | |
| δ (біреу) |
Голофермент РНҚ- полимеразаның құрылымы былай жазылады: 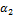 ββ’ωδ. Ол екі компоненттен тұрады: минимальдық ферменттен ( ββ’ω) және сигма-фактордан (δ суббөлік). Олардың қызметі әр түрлі: тек қана голофермент транскрипцияны бастай алады, онан кейін сигма-фактор комплекстен бөлініп шығады, ал РНҚ элонгациясын минимальдық фермент жалғастырады. Ол нуклеотидтер арасында фосфодиэфир байланыстарының түзілуін катализдейді, бірақ керекті жерінде транскрипцияны бастай алмайды. Ферменттің активті орталығына
ββ’ωδ. Ол екі компоненттен тұрады: минимальдық ферменттен ( ββ’ω) және сигма-фактордан (δ суббөлік). Олардың қызметі әр түрлі: тек қана голофермент транскрипцияны бастай алады, онан кейін сигма-фактор комплекстен бөлініп шығады, ал РНҚ элонгациясын минимальдық фермент жалғастырады. Ол нуклеотидтер арасында фосфодиэфир байланыстарының түзілуін катализдейді, бірақ керекті жерінде транскрипцияны бастай алмайды. Ферменттің активті орталығына 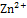 кіреді.
кіреді.
Эукариот РНҚ полимеразалары күрделі құралған, әлі толық зерттелмеген. РНҚ-полимеразалар көптеген пептидтік тізбектерден тұрады. Полимеразалық активтігі бар тазартылмаған фермент препараттары 500 000 дейін молекулалық массасы бар үлкен белоктар, әрбір РНҚ-полимераза 2 суббөлікткен тұрады, олардың М 200 000 және 140 000.
Олардан басқа он шақты кіші суббөліктер табылған, М 10 000-нан 90 000 дейін. Әлі бірде-бір РНҚ-полимераза таза түрінде алынбағандықтан, олардың құрамы мен құрылымы туралы сипаттама беру қиын.
РНҚ-полимераза I негізінде ядрошықта орналасқан, рРНҚ гендерінің транскрипциясына жауапты. Ол РНҚ-лар 50-70% мөлшерінің синтезін қамтамасыз етеді, магний немесе марганец иондарымен активтенеді.
РНҚ-полимераза II нуклеоплазмада табылған. Ол мРНҚ ізашары гетерогенді РНҚ синтезіне жауапты., оның үлесіне клеткада түзілетін РНҚ-лардың 20-40% тиеді. Магний иондарымен активтенеді.
РНҚ-полимераза III нуклеоплазмада орналасқан, көптеген кіші РНҚ және тРНҚ минтезін катализдейді, клеткадағы РНҚ-ның 10% осының үлесіне келеді. Марганец иондарымен активтенеді.
2. Прокариоттардағы транскрипция процесінің реттелу механизмі.
Оперон — геномның бөлігі, ондағы мәлімет иРНҚ бір молекуласына репрессор белоктың бақылауымен көшіріледі. Оперон реттеуі (транскрипция деңгейіндегі реттеу)прокариоттар және бактериофагтар гендерінің белсенділігін реттеудегі негізгі механизм. Оперон бір-бірімен тығыз байланысқан бірнеше құрылымдық гендерден тұрады. Бұл гендер бір метаболиттің биосинтез сатыларына қатынасатын белоктардың коды болып табылады. Сонымен бірге оперон өзі реттеуші кұрамдардан тұрады: оперон алдында — промотор (транскрипцияның бастапкы белігі) және оператор (белоксупрессордың байланысатын бөлігі), оперон соңында — терминатор (транскрипцияның тоқтайтын бөлігі).
Реттеуші орталық құрамына ген кіреді және әруақытта жұмыс халінде болады және экспрессия спецификалық реттелуге ұшырамайды, сондықтан конститутивті деп аталынады. Бұл геннің өн

