 2015-07-04
2015-07-04 1284
1284Рецепторы соматосенсорной системы, в соответствии с типом стимула, на который они отвечают, можно разделить на три основных типа: механорецепторы, терморецепторы и ноцицепторы.
Механорецепторы кожи реагируют на тактильные стимулы – прикосновение к коже, поглаживание или давление. Большинство их них являются инкапсулированными нервными окончаниями, то есть имеют специализированную капсулу, опосредующую влияние механической деформации на мембрану нервной терминали. Выделяют быстро адаптирующиеся – волосяные рецепторы, тельца Мейсснера, тельца Пачини, а
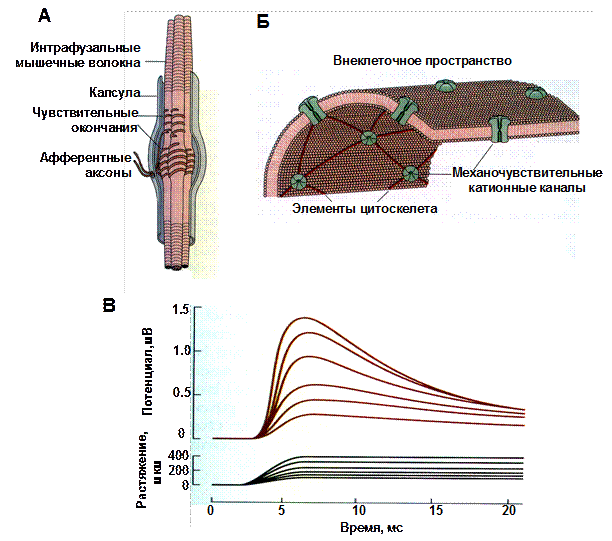
Рис. 44. Механорецепция в мышечных веретенах
На верхней части рисунка показаны строение мышечного веретена (А) и механочувствительной мембраны сенсорных окончаний (Б). В – ответы изолированного мышечного веретена на растяжение. Верхняя запись показывает рецепторные потенциалы, возникающие в сенсорных окончаниях при растяжении на разную длину. Нижняя запись отражает амплитуду растяжений. Начальная деполяризация чувствительных окончаний пропорциональна амплитуде и скорости растяжения (динамический ответ), однако, когда растяжение фиксируется на определенной длине, рецепторный потенциал спадает до более низких значений, пропорциональных только амплитуде растяжения (статический ответ).
также медленно адаптирующиеся – диски Меркеля и тельца Руффини. При этом наиболее поверхностно из них расположены тельца Мейсснера и диски Меркеля, а глубоко – тельца Пачини и Руффини (Рис. 45). Механорецепторы иннервируются толстыми миелинизированными волокнами, проводящими потенциалы действия с большой скоростью.
Трансдукция в механорецепторах происходит благодаря наличию механочувствительных ионных каналов в мембране нервного окончания, при активации которых возникает входящий катионный (натриевый или кальциевый) ток, обеспечивающий возникновение деполяризующего рецепторного потенциала с последующей генерацией потенциала действия. В отсутствии тактильных стимулов механорецепторы «молчат».
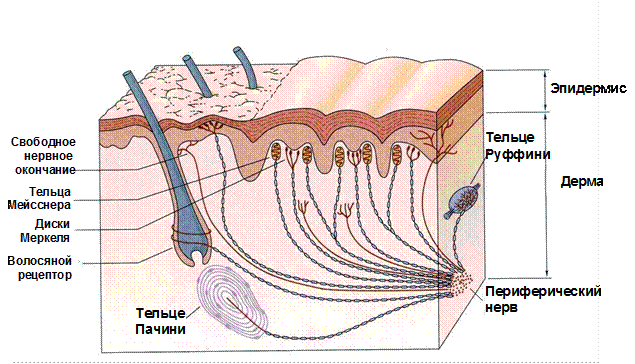
Рис. 45. Локализация и морфология механорецепторов кожи
Многие механорецепторы располагаются поверхностно, в области границы эпидермиса и дермы (свободные нервные окончания, тельца Мейсснера, диски Меркеля), тогда как другие – более глубоко, в дерме и подкожной клетчатке (тельца Пачини и Руффини). Рецепторный аппарат кожи без волосяного покрова (пальцев, ладоней, губ и др.) и волосистой кожи несколько различен. Например, тельца Мейсснера располагаются преимущественно в безволосой коже, тогда как в волосистой коже расположено больше свободных нервных окончаний.
Терморецепторы, обеспечивающие температурную чувствительность кожи, представляют собой свободные нервные окончания. Термальные ощущения происходят из разности между температурой объекта, контактирующего с кожей, и нормальной температурой кожи (340 С). В отличие от механорецепторов, тепловые и холодовые рецепторы даже при нормальной температуре с небольшой частотой генерируют потенциалы действия (2-5 имп/с). При охлаждении исчезает импульсация тепловых рецепторов, а при нагревании – холодовых. Максимальная активность холодовых рецепторов наблюдается при 250 С, а тепловых – при 450С. Дальнейшее снижение или повышение температуры приводит к снижению активности терморецепторов. Из этого следует, что во-первых, они не могут сигнализировать о боли, связанной с сильным нагреванием или охлаждением, а во-вторых, один только тепловой или холодовой рецептор не может дать точную информацию о температуре.
Последние исследования показали, что термотрансдукция связана с наличием специальных, температуро-чувствительных ионных каналов из семейства TRP (transient receptor potential – «временный рецепторный потенциал») на мембранах терморецепторов. Эти каналы проницаемы для катионов, особенно для кальция. На данный момент существует несколько разновидностей температуро-чувствительных TRP-каналов (TRPV1, TRPV3, TRPM8, TRPA1), активирующихся при различных значениях температуры; этот ряд продолжает расти.
Таким образом, изменение температуры приводит к активации TRP-каналов мембраны рецептора, появлению (или возрастанию) входящего катионного тока и увеличению частоты генерации потенциалов действия. Возбуждение от терморецепторов проводится по Аδ и С-волокнам.
Болевые рецепторы, или ноцицепторы реагируют на стимулы, угрожающие организму повреждением. Выделяют Аδ-механорецепторы и полимодальные С-ноцицепторы. Аδ-механорецепторы реагируют на сильные механические воздействия, такие как, например, укол иглой. С-ноцицепторы реагируют на стимулы разной модальности – химические, механические, температурные (выше 450С или ниже 50С). Известно, что боль вызывают многие химические факторы - гистамин, брадикинин, высвобождение калия из поврежденных клеток, АТФ, серотонин, вещество P, повышение кислотности, ацетилхолин и др. Это свидетельствует о том, что многие болевые рецепторы являются хеморецепторами.
Поскольку боль вызывают стимулы различных модальностей, существует довольно много трансдукционных механизмов боли, многие из которых изучены недостаточно.
Важную роль в трансдукции боли играют катионные каналы из уже упомянутого нами семейства TRP. Наиболее изученным из них является TRPV1-канал, или «ванилоидный рецептор», который чувствителен к нескольким воздействиям - повышенной температуре (порог активации около 430C), капсаицину – компоненту «чилийского» жгучего перца и повышенной кислотности. Существует другие – TRPV2-каналы, которые за счет чувствительности к более высокой температуре (530C) обеспечивают болевую рецепцию избыточного нагревания.
Что касается детекции болевых механических стимулов, то на данный момент открыто два механизма. Первый связан с наличием в мембране рецепторов механочувствительных катионных каналов из семейства дегенеринов. Второй механизм, показанный на ряде объектов, связан с увеличением концентрации внеклеточной АТФ в тканях при их деформации. Внеклеточная АТФ активирует G-белок-связанные АТФ-чувствительные ионные каналы на мембране ноцицепторов, что приводит к деполяризации и генерации потенциалов действия.
Итак, любое сенсорное восприятие начинается с возникновения градуального электрического сигнала – рецепторного потенциала на мембране воспринимающей клетки. Чем сильнее и дольше стимул будет действовать на организм, тем больше по амплитуде и длительнее будет рецепторный потенциал. В конечном итоге возникновение рецепторного потенциала будет влиять на частоту и количество потенциалов действия, генерируемых и проводящихся чувствительным нейроном в центральную нервную систему. В случае деполяризационного рецепторного потенциала эти параметры будут возрастать, а в случае гиперполяризационного – снижаться. Следовательно, любой сенсорный сигнал попадает в мозг в виде частотно-импульсного кода, то есть последовательности некоторого количества потенциалов действия, идущих с определенной частотой. Любое сенсорное ощущение при этом будет определяться типом и количеством активированных рецепторов, частотой и количеством потенциалов действия, идущих по афферентным нервам от рецепторов, а также тем, в какие участки мозга эти сигналы попадают.
Как вы помните, нейроны, получая информацию от соседних нервных клеток в виде постсинаптических потенциалов на своей мембране, обрабатывают и передают ее другим клеткам также в виде частотно-импульсного кода потенциалов действия. Именно так мотонейроны «принимают» решение – совершать организму движение или нет. Таким образом, этот универсальный механизм кодирования информации, основанный на интеграции градуальных электрических сигналов на мембране клетки и генерации неградуальных – потенциалов действия, лежит в основе всех функций нервной системы.
Таким образом, современная физиология далеко продвинулась в понимании электрической сигнализации возбудимых клеток. Тем не менее, многие вопросы пока остаются непонятными. Несомненно, дальнейшее изучение процессов, лежащих в основе электрических событий на мембранах возбудимых клеток, позволит нам приблизиться к пониманию высших интегративных функций мозга и организма в целом. Завершая данное пособие, нам бы хотелось привести высказывание выдающегося современного нейробиолога Эрика Кэндела, получившего в 2000 году совместно с Арвидом Карлссоном и Полом Грингардом Нобелевскую премию «За открытие механизмов передачи сигналов в нервной системе» (рис. 46):
«Прогресс в понимании основных умственных функций происходит благодаря подходу, основанному на том допущении, что эти функции возникают из биологических свойств нервных клеток и манеры их взаимодействий. Согласно этому взгляду, разум может быть рассмотрен как набор операций, осуществляемых мозгом – органом, обрабатывающим информацию, который обладает большой мощностью благодаря огромному количеству, разнообразию, взаимодействиям его нервных клеток и сложности контактов между ними».








