 2015-09-06
2015-09-06 375
375БИОМАКРОМОЛЕКУЛАХ [30]
Прошло несколько десятилетий после того, как лауреаты Нобелевской премии академики РАН А.Н.Прохоров, Н.Г.Басов (Россия) и Чарльз Таунс (США), высказали идею, а затем реализовали ее, о возможности создания квантовых генераторов. Сейчас трудно сказать, в какой области науки и техники они не применяются (от биологии и медицины до лазерного термоядерного синтеза). Последующие исследователи внесли свой крупный вклад в развитие этой проблемы.
В данной части работы ставится вопрос: можно ли in vitro создать лазер на информационных биомакромолекулах, прежде всего на ДНК, РНК и хромосомах? Вряд ли может идти речь о создании энергетически мощных лазеров на этих структурах. Вопрос звучит по-иному: какие новые знания мы можем получить о ДНК, РНК и хромосомах, создав такой лазер и исследуя характер его излучения? Можно думать, что это будут принципиально новые данные. Например, об их нелинейной динамике, в том числе солитонного типа, о ровибронных колебаниях, о модуляциях дисперсии оптического вращения и кругового дихроизма, переносе энергии в другие, ранее недоступные (в таком варианте методологии) слои информации. При этом динамические модификации лазерного пучка могут иметь cемантико-гено-биознаковый характер и поэтому будут обладать мощной биологической активностью.
Первые соображения по этому поводу были предложены нами ранее [25,30]. В том числе обсуждалась идея о создании лазерной системы на Фрёлиховских модах [3]. Сложность доказательства правильности всех этих мыслей состоит в том, что большинство генетических структур, содержащих в своем составе ароматические и гетероциклические кольца, “прозрачны” для характерного спектрального диапазона l@350400нм. Трудность также и в том, что если использовать мощную оптическую накачку, то это, учитывая “хрупкость” биоструктур, неизбежно приведёт к их разрушению.
В настоящей главе для реализации некоторых из обсуждавшихся положений проведено исследование in vitro спектров двухфотонно-возбуждаемой люминесценции (ДВЛ) геле-жидкокристаллических препаратов нуклеогистона, являющегося суммарной фракцией хромосом, в которой преобладают гистоновые белки, и ДНК (стандартные высокополимерные препараты фирмы “Sigma”). Для существенного увеличечения интенсивности ДВЛ генетических структур нами предложен способ активации люминесценции за счет введения в состав исследуемых образцов активаторов (доноров) ДВЛопределенных (близких по спектру оптического поглощения ДНК и нуклеогистону) органических молекул. Такие молекулы характеризуются большой интенсивностью спектров излучения, которые располагаются в области собственного оптического поглощения ДНК и нуклеогистона. В качестве активатора мы использовали кристаллический препарат димедрола, структура которого включает пару бензольных колец. Для димедрола это обеспечивает интенсивный спектр ДВЛ, имеющий вид широкой асимметричной полосы в диапазне 280 350нм.
Для фотонной накачки исследуемых препаратов мы применяли лазер на парах меди. Этот лазер работает в стандартном импульсно-периодическом режиме с частотой следования импульсов 10кГц, со средней мощностью 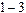 Вт, пиковой мощностью 10
Вт, пиковой мощностью 10 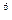 Вт, длинами волн генерации l = 510,8нм и 578,2нм (зеленая и желтая линии), длительностью импульсов
Вт, длинами волн генерации l = 510,8нм и 578,2нм (зеленая и желтая линии), длительностью импульсов 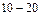 нс. Лазерное излучение направляли на исследуемый образец в виде сфокусированного пятна размером
нс. Лазерное излучение направляли на исследуемый образец в виде сфокусированного пятна размером 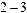 мм. Применение такого лазера как инициатора ДВЛ оказалось весьма эффективным при изучении электронно-колебательных спектров белков, ДНК, нуклеогистона и их компонентов (пурины, пиримидины, аминокислоты [19,30]). Регистрирующая аппаратура включала: фильтр для выделения лазерных линий с l=510,8 и 578,2 нм, фильтр для выделения излучений люминесценции в УФ и фиолетовом диапазонах (с подавлением лазерного излучения), монохроматор (тип МДР2) для сканирования спектра в широком интервале (от УФ до видимой области), двухкоординатный самописец для регистрации спектров, измеритель для контроля опорного сигнала и определения эффективности наблюдаемого сигнала. Для подавления тепловых шумов применяли строб-импульс длительностью 2530нс, синхронизированный с импульсом возбуждения. Регистрацию вторичного импульса излучения проводили с ФЭУ130. Исследования спектров ДВЛ геле-жидкокристаллического препарата ДНК в смеси с димедролом (ДНК-ДЛ) и нуклеогистона с димедролом (НГ-ДЛ) показали, что амплитуда ДВЛ спектра ДНК-ДЛ лишь на порядок меньше таковой спектра ДВЛ чистого димедрола. Это обеспечивает существенное увеличение интенсивности ДВЛ смеси ДНК-ДЛ по сравнению с чистым препаратом ДНК в форме жесткого геля [19]. На этом же спектре обнаруживается ряд дополнительных особенностей изучаемых смесей. Оказалось, что квантовый выход ДВЛ для смеси НГ-ДЛ ниже, чем для смеси ДНК-ДЛ. Другая характерная черта разгорание или тушение ДВЛ во времени. Для НГ-ДЛ наблюдается нарастание ДВЛ во времени. Обратный эффект наблюдается в случае ДНК-ДЛ. Представляет интерес присутствие вибронной структуры в спектрах ДВЛ в виде отдельных перекрывающихся полос в области 310370 нм, особенно для ДНК-ДЛ. Такая структура близка к ранее наблюдавшимся спектрам ДВЛ для нуклеозид-трифосфатов [19].
мм. Применение такого лазера как инициатора ДВЛ оказалось весьма эффективным при изучении электронно-колебательных спектров белков, ДНК, нуклеогистона и их компонентов (пурины, пиримидины, аминокислоты [19,30]). Регистрирующая аппаратура включала: фильтр для выделения лазерных линий с l=510,8 и 578,2 нм, фильтр для выделения излучений люминесценции в УФ и фиолетовом диапазонах (с подавлением лазерного излучения), монохроматор (тип МДР2) для сканирования спектра в широком интервале (от УФ до видимой области), двухкоординатный самописец для регистрации спектров, измеритель для контроля опорного сигнала и определения эффективности наблюдаемого сигнала. Для подавления тепловых шумов применяли строб-импульс длительностью 2530нс, синхронизированный с импульсом возбуждения. Регистрацию вторичного импульса излучения проводили с ФЭУ130. Исследования спектров ДВЛ геле-жидкокристаллического препарата ДНК в смеси с димедролом (ДНК-ДЛ) и нуклеогистона с димедролом (НГ-ДЛ) показали, что амплитуда ДВЛ спектра ДНК-ДЛ лишь на порядок меньше таковой спектра ДВЛ чистого димедрола. Это обеспечивает существенное увеличение интенсивности ДВЛ смеси ДНК-ДЛ по сравнению с чистым препаратом ДНК в форме жесткого геля [19]. На этом же спектре обнаруживается ряд дополнительных особенностей изучаемых смесей. Оказалось, что квантовый выход ДВЛ для смеси НГ-ДЛ ниже, чем для смеси ДНК-ДЛ. Другая характерная черта разгорание или тушение ДВЛ во времени. Для НГ-ДЛ наблюдается нарастание ДВЛ во времени. Обратный эффект наблюдается в случае ДНК-ДЛ. Представляет интерес присутствие вибронной структуры в спектрах ДВЛ в виде отдельных перекрывающихся полос в области 310370 нм, особенно для ДНК-ДЛ. Такая структура близка к ранее наблюдавшимся спектрам ДВЛ для нуклеозид-трифосфатов [19].
Механизм резкого увеличения квантового выхода ДВЛ нуклеогистона и ДНК при наличии донор-активатора (димедрола) может быть объяснен быстрой квазирезонансной передачей энергии от возбужденных молекул димедрола к исследуемым геноструктурам. Наблюдаемая при этом тонкая многополосчатая структура ДВЛ спектров коррелирует с характером вибронных полос для ряда ароматических и гетероциклических соединений, включая чистые нуклеозид-трифосфаты и ДНК [19]. Возникновение такого рода дискретизации спектров можно трактовать переходом электронов биомакромолекул с электронного терма S1 на возбужденные колебательные уровни основного состояния S0. В связи с этим может быть реализована инверсная заселенность на переходах 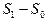 при достаточном заселении терма
при достаточном заселении терма 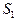 .
.
Проведем оценки необходимой интенсивности 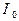 лазерного излучения для создания инверсии (суперфлуоресценции) в условиях проведенных опытов.
лазерного излучения для создания инверсии (суперфлуоресценции) в условиях проведенных опытов.
Условия инверсии записываются следующим образом:
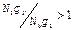 , (1)
, (1)
где 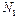 - плотность рабочих молекул в состоянии ,
- плотность рабочих молекул в состоянии , 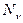 - плотность молекул в состоянии
- плотность молекул в состоянии 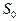 ,
, 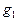 и
и 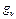 - соответствующие статистические веса квантовых уровней.
- соответствующие статистические веса квантовых уровней.
Плотность заселенности оценивается из соотношения
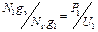 , (2)
, (2)
где 
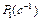 ‑ скорость заселения уровня
‑ скорость заселения уровня 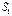 ,
, 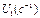 - скорость его распада за счет излучательного процесса и (или) безызлучательных процессов.
- скорость его распада за счет излучательного процесса и (или) безызлучательных процессов.
Для величины 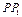 имеем оценку:
имеем оценку:
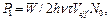 (3)
(3)
где W и 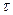 - энергия и длительность лазерного импульса,
- энергия и длительность лазерного импульса, 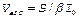 - эффективный объем среды, в котором реализуется двухфотонное поглощение (S - площадь поперечного сечения сфокусированного светового пучка, падающего на исследуемый образец,
- эффективный объем среды, в котором реализуется двухфотонное поглощение (S - площадь поперечного сечения сфокусированного светового пучка, падающего на исследуемый образец, 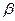 - эффективная длина проникновения излучения в образец),
- эффективная длина проникновения излучения в образец), 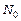 - плотность биомакромолекул (ДНК или нуклеогистона).
- плотность биомакромолекул (ДНК или нуклеогистона).
С учетом соотношений (1) (3) условие для cоздания инверсной заселённости суперфлуоресценции записывается в виде
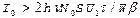 .
.
Используя характерные данные
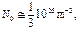
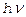 (для
(для 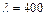 нм) =
нм) = 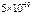 Дж,
Дж,
t@ 10нс, 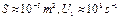 ,
, 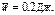
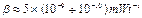 ,
,
получаем оценку 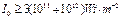 ,
,
что близко к использованным значениям интенсивности в наших экспериментах.
Проведенные экспериментальные исследования и их теоретические оценки дают основание достаточно уверенно предполагать, что при используемых режимах двухфотонного возбуждения с использованием активатора-димедрола в геноструктурах in vitro реализуется усиление люминесценции, т.е. излучение ДНК и нуклеогистона носит характер суперфлуоресценции.
Не исключено, что в биосистеме роль димедролоподобных веществ в качестве активаторов могут выполнять эндогенные соединения, прямо или косвенно взаимодействующие с ДНК и хромосомами (стероидные гормоны, углеводы, нуклеозид -моно, -ди и -трифосфаты, некоторые витамины (например, рибофлавин), ароматические и гетероциклические аминокислоты, катехол- и индолалкиламины, некоторые антибиотики, наркотические вещества (например, эндогенные морфины метаболиты этанола и пептиды-эндорфины), алкалоиды, токсины, ко-факторы ферментов, гем-содержащие белки и другие многочисленные органические соединения, содержащие бензольные и гетероциклические компоненты.
Неясны условия реализации инверсной электронной заселенности геноструктур in vivo, близкие тем, которые использовались нами в режимах ДВЛ. Такие условия могут создаваться в биосистемах, например, за счет фотон-фононных взаимодействий в ДНК в рамках теории Дике[28].
Однако, это относится к чисто физическим механизмам. Что касается физико-биохимических процессов, приводящих к лазерной накачке ДНК и хромосом in vivo, то в качестве таковых можно предсказать наличие в биосистемах мощных АТФ-азных систем, поставляющих энергию для перевода генетических структур в биокогерентные состояния (аналогичные тем, что как частный случай изложены в настоящей главе).








