 2014-02-17
2014-02-17 2694
2694Регуляция биосинтеза белка
В каждой клетке синтезируются специфические белки, и с неодинаковой скоростью. Благодаря регуляции синтеза в конкретных условиях среды образуется лишь необходимое число молекул данного белка. Все соматические клетки многоклеточных организмов содержат в ДНК одинаковую генетическую информацию, однако отличаются друг от друга по составу белков. Так, клетки эритроцитов содержат большое количество гемоглобина, клетки кожи – коллагена, скелетных мышц – актина и миозина, клетки печени содержат ферменты синтеза мочевины, которые отсутствуют у всех других клеток. Таким образом, в клетках каждого типа экспрессируется только часть структурных генов.
Большая часть генома находится в неактивном, репрессированном, состоянии. Спектр функционирующих генов зависит от типа клетки, периода ее жизненного цикла, стадии индивидуального развития организма. У большинства организмов активно транскрибируются только 2-10% генов.
Гены, которые транскрибируются постоянно, не подчиняясь каким-либо регуляторным воздействиям, называются конститутивными. Обычно это гены, обеспечивающие синтез белков общего назначения (белки рибосом, гистоны, тубулины и др.), а также тРНК и рРНК. Включение и выключение других генов зависит от различных метаболитов, эти гены называются регулируемыми.
Схема регуляции активности генов на уровне транскрипции была впервые разработана Ф. Жакобом и Ж. Моно (1961) на примере лактозного (lac)-оперона кишечной палочки (E.coli). Единица регуляции транскрипции была названа опероном. Оперон – это последовательность структурных генов, определяющих синтез группы белков, участвующих в одной метаболической цепи, имеющих общий промотор и оператор.
E.coli быстро растет на питательной среде, содержащей в качестве источника углерода глюкозу. Глюкоза – самый доступный и выгодный для E.coli источник углерода и энергии. Пока глюкоза имеется в достаточном количестве, клетка не нуждается в получении и переработке других сахаров.
После переноса клеток на среду, содержащую вместо глюкозы лактозу, рост сначала замедляется, а затем возобновляется с высокой скоростью. При этом бактерии синтезируют три необходимых для усвоения лактозы фермента:
· β-галактозидазу, которая расщепляет лактозу на глюкозу и галактозу;
· пермеазу, необходимую для проникновения лактозы через клеточную мембрану;
· тиогалактозидазу.
Эти ферменты кодируются тремя структурными генами (соответственно Z, Y и А) лактозного оперона. Кроме структурных генов в состав оперона входят промотор и оператор. В схеме оперонной регуляции присутствует также ген-регулятор, кодирующий белок-репрессор. Однако ген-регулятор не входит в состав оперона и может находиться на расстоянии от него.
При отсутствии в среде лактозы активный белок-репрессор связывается с оператором, блокируя транскрипцию структурных генов. Хотя РНК-полимераза может связаться с промотором, она не перемещается дальше «выключенного» оператора (рис. 45А). При добавлении в среду лактозы ее молекулы связываются со специфическим участком репрессора, инактивируя его. У инактивированного репрессора резко снижается сродство к зоне оператора, в результате чего он отсоединяется, и РНК-полимераза начинает транскрипцию структурных генов (рис. 45Б).
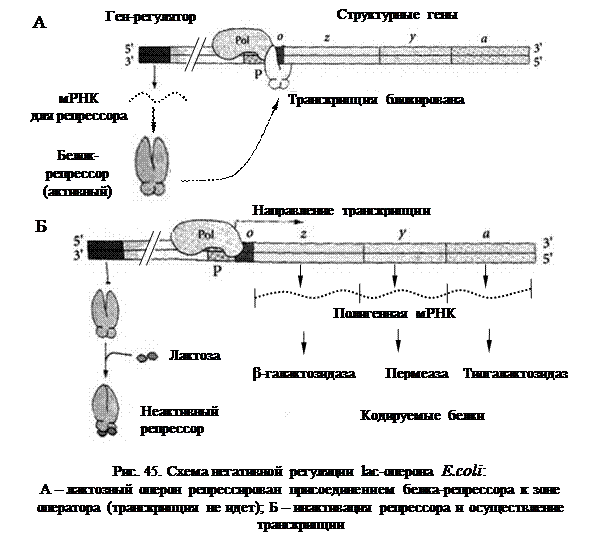
Снижение концентрации субстрата – лактозы – вновь является сигналом к соединению белка-репрессора с оператором и прекращению транскрипции генов lac-оперона. Таким образом, синтез ферментов происходит только в том случае, если в них возникает необходимость. Данная схема регуляции называется «негативной», так как контролирующим транскрипцию фактором является негативный фактор, «выключатель» – белок-репрессор. Индукция (включение) происходит при потере сродства белка-репрессора к оператору.
Наряду с негативной системой регуляции, для lac -оперона существует и позитивная регуляция. Промотор лактозного оперона способен связывать не только РНК-полимеразу, но и особый белок-активатор катаболизма (САР) в комплексе с циклическим АМФ (рис. 46А).
Присутствие САР и цАМФ вызывает не репрессию, а напротив, активирование транскрипции. Без САР РНК-полимераза не может связаться с промотором и начать транскрипцию. САР, образовав комплекс с цАМФ,
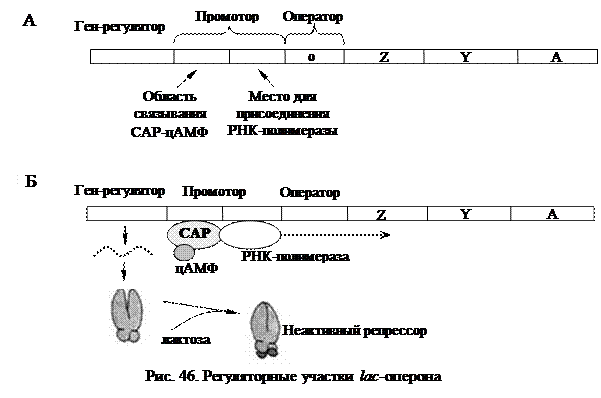
активизируется и только после этого присоединяется к своему сайту (САР-участку) на промоторе, многократно (почти в 50 раз) усиливая транскрипцию генов lac -оперона. При этом транскрипция возможна только в присутствии лактозы, когда оператор не блокирован репрессором (рис. 46Б). В случае присутствия в среде глюкозы, концентрация цАМФ в клетке резко снижается, и не образуется комплекса цАМФ с САР. В результате этого РНК-полимераза не может связаться с промотором и lac-гены не транскрибируются.

