 2015-05-13
2015-05-13 12294
12294Белки. Структура белков. Свойства белков.
Белок – это последовательность аминокислот, связанных друг с другом пептидными связями.
Легко представить, что количество аминокислот может быть различно: от минимум двух до любых разумных величин. Биохимики условились считать, что если количество аминокислот не превышает 10, то такое соединение называется пептид; если от 10 и более аминокислот – полипептид. Полипептиды, способные самопроизвольно формировать и удерживать определенную пространственную структуру, которая называется конформацией, относят к белкам. Стабилизация такой структуры возможна лишь при достижении полипептидами определенной длины (более 40 аминокислот), поэтому белками обычно считают полипептиды молекулярной массой более 5 000 Да. (1Да равен 1/12 изотопа углерода). Только имея определенное пространственное строение (нативную структуру), белок может выполнять свои функции.
Размер белка может измеряться в дальтонах (молекулярная масса), чаще из-за относительно большой величины молекулы в производных единицах — килодальтонах (кДа). Белки дрожжей, в среднем, состоят из 466 аминокислот и имеют молекулярную массу 53 кДа. Самый большой из известных в настоящее время белков — титин — является компонентом саркомеров мускулов; молекулярная масса его различных изоформ варьирует в интервале от 3000 до 3700 кДа, он состоит из 38 138 аминокислот (в человеческой мышце solius).
Структура белка
Трехмерная структура белка формируется в процессе фолдинга (от англ. folding - «сворачивание»). Трехмерная структура формируется в результате взаимодействия структур более низких уровней.
Выделяют четыре уровня структуры белка:
Первичная структура — последовательность аминокислот в полипептидной цепи.
Вторичная структура — это размещение в пространстве отдельных участков полипептидной цепи.
Ниже приведены самые распространенные типы вторичной структуры белков:
α-спирали — плотные витки вокруг длинной оси молекулы, один виток составляют 3,6 аминокислотных остатка, и шаг спирали составляет 0,54 нм (на один аминокислотный остаток приходится 0,15 нм), спираль стабилизирована водородными связями между H и O пептидных групп, отстоящих друг от друга на 4 аминокислотных остатка. Спираль построена исключительно из одного типа стереоизомеров аминокислот (L). Хотя она может быть как левозакрученной, так и правозакрученной, в белках преобладает правозакрученная. Спираль нарушают электростатические взаимодействия глутаминовой кислоты, лизина, аргинина. Расположенные близко друг к другу остатки аспарагина, серина, треонина и лейцина могут стерически мешать образованию спирали, остатки пролина вызывает изгиб цепи и также нарушают структуру α-спирали.
β-складчатые слои — несколько зигзагообразных полипептидных цепей, в которых водородные связи образуются между относительно удаленными друг от друга (0,347 нм на аминокислотный остаток) в первичной структуре аминокислотами или разными цепями белка, а не близко расположенными, как имеет место в α-спирали. Эти цепи обычно направлены N-концами в противоположные стороны (антипараллельная ориентация). Для образования β-складчатых слоев важны небольшие размеры боковых групп аминокислот, обычно преобладают глицин и аланин.
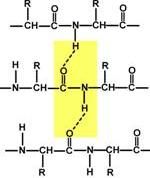 Участие водородных связей в формировании вторичной структуры Участие водородных связей в формировании вторичной структуры | 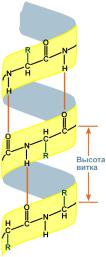 Укладка белка в виде α-спирали Укладка белка в виде α-спирали |
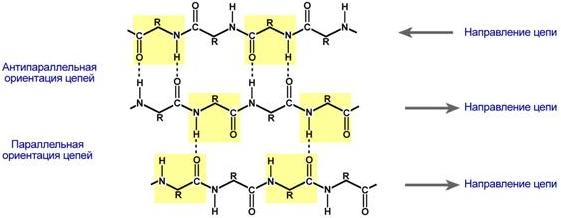
Укладка белка в виде β-складчатого слоя
Неупорядоченные структуры - это неупорядоченное расположение белковой цепи в пространстве.
Пространственная структура каждого белка индивидуальна и определяется его первичной структурой. Однако сравнение конформаций разных по структуре и функциям белков выявило наличие у них похожих сочетаний элементов вторичной структуры. Такой специфический порядок формирования вторичных структур называют супервторичной структурой белков. Супервторичная структура формируется за счёт межрадикальных взаимодействий.
Определённые характерные сочетания α-спиралей и β-структур часто обозначают как "структурные мотивы". Они имеют специфические названия: "α-спираль-поворот-α-спираль", "структура α/β-бочонка", "лейциновая застёжка-молния", "цинковый палец" и др.
Третичная структура – это способ размещения в пространстве всей полипептидной цепи. Наряду с α-спиралями, β-складчатыми слоями и супервторичными структурами в третичной структуре обнаруживается неупорядоченная конформация, которая может занимать значительную часть молекулы.
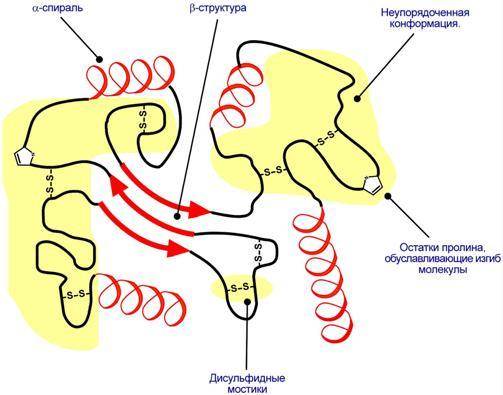
Схематичное представление укладки белка в третичную структуру.
Четвертичная структура возникает у белков, которые состоят из нескольких полипептидных цепей (субъединиц, протомеров или мономеров), при объединении третичных структур этих субъдиниц. Например, молекула гемоглобина состоит из 4 субъединиц. Четвертичную структуру имеют надмолекулярные образования – мультиферментные комплексы, которые состоят из нескольких молекул ферментов и коферментов (пируватдегидрогеназа), и изоферменты (лактатдегидрогеназа - ЛДГ, креатинфосфокиназа – КФК).
Итак. Пространственная структура зависит не от длины полипептидной цепи, а от последовательности аминокислотных остатков, специфичной для каждого белка, а также от боковых радикалов, свойственных соответствующим аминокислотам. Пространственную трехмерную структуру или конформацию белковых макромолекул образуют в первую очередь водородные связи, гидрофобные взаимодействия между неполярными боковыми радикалами аминокислот, ионные взаимодействия между противоположно заряженными боковыми группами аминокислотных остатков. Водородные связи играют огромную роль в формировании и поддержании пространственной структуры белковой макромолекулы.
Что касается гидрофобных взаимодействий, то они возникают в результате контакта между неполярными радикалами, неспособными разорвать водородные связи между молекулами воды, которая вытесняется на поверхность белковой глобулы. По мере синтеза белка неполярные химические группировки собираются внутри глобулы, а полярные вытесняются на ее поверхность. Таким образом, белковая молекула может быть нейтральной, заряженной положительно или же отрицательно в зависимости от рН растворителя и ионогенных групп в белке. Кроме того, конформация белков поддерживается ковалентными связями S—S, образующимися между двумя остатками цистеина. В результате образования нативной структуры белка многие атомы, находящиеся на удаленных участках полипептидной цепи, сближаются и, воздействуя друг на друга, приобретают новые свойства, отсутствующие у индивидуальных аминокислот или небольших полипептидов.
Важно понимать, что фолдинг — сворачивание белков (и других биомакромолекул) из развёрнутой конформации в «нативную» форму — физико-химический процесс, в результате которого белки в своей естественной «среде обитания» (растворе, цитоплазме или мембране) приобретают характерные только для них пространственную укладку и функции.
В клетках имеется ряд каталитически неактивных белков, которые тем не менее вносят большой вклад в образование пространственных структур белков. Это так называемые шапероны. Шапероны помогают правильной сборке трехмерной белковой конформации путем образования обратимых нековалентных комплексов с частично свернутой полипептидной цепью, одновременно ингибируя неправильно образованные связи, ведущие к формированию функционально неактивных белковых структур. В перечень функций, свойственных шаперонам, входит защита расплавленных (частично свернутых) глобул от агрегации, а также перенос новосинтезированных белков в различные локусы клеток.
Шапероны преимущественно являются белками теплового шока, синтез которых резко усиливается при стрессовом температурном воздействии, поэтому их называют еще hsp (heat shock proteins). Семейства этих белков найдены в микробных, растительных и животных клетках. Классификация шаперонов основана на их молекулярной массе, которая варьирует от 10 до 90 kDa. Они являются белками-помощниками процессов образования трехмерной структуры белков. Шапероны удерживают новосинтезированную полипептидную цепь в развернутом состоянии, не давая ей свернуться в отличную от нативной форму, и обеспечивают условия для единственно правильной, нативной структуры белка.
В процессе фолдинга белка некоторые конформации молекулы забраковываются на стадии расплавленной глобулы. Деградацию таких молекул инициирует белок убиквитин.
Деградация белка по убиквитиновому пути включает две основные стадии:
1) ковалентное присоединение убиквитина к подлежащему деградации белку через остаток лизина, наличие такой метки в белке является первичным сигналом сортировки, направляющей образовавшиеся конъюгаты к протеасомам, в большинстве случаев к белку присоединяется несколько молекул убиквитина, которые организованы в виде бусинок на нитке.;
2) гидролиз белка протеосомой (основная функция протеасомы – протеолитическая деградация ненужных и повреждённых белков до коротких пептидов). Убиквитин заслуженно называют «меткой смерти» для белка.
Доме?н белка? – элемент третичной структуры белка, представляющий собой достаточно стабильную и независимую подструктуру белка, чей фолдинг проходит независимо от остальных частей. В состав домена обычно входит несколько элементов вторичной структуры. Сходные по структуре домены встречаются не только в родственных белках (например, в гемоглобинах разных животных), но и в совершенно разных белках. Белок может иметь несколько доменов, эти участки могут выполнять разные функции в одном и том же белке. Доменную структуру имеют некоторые ферменты и все иммуноглобулины. Белки с длинными полипептидными цепями (более 200 аминокислотных остатков) часто создают доменные структуры.

