 2015-05-25
2015-05-25 1581
1581В шестидесятых годах в нейрофизиологической литературе стали появляться отдельные сообщения о том, что многократная, производимая с достаточной частотой стимуляция нервных путей к некоторым областям коры головного мозга приводит к длительному усилению спонтанной электрической активности таких областей. Это явление - в сущности, повышение эффективности передачи между пре- и постсинаптическими клетками - получило название потенциации. Не может ли такая потенциация быть одной из форм нейрофизиологической памяти? В 1973 году в очень часто цитируемой теперь статье Тим Блисс из Национального института медицинских исследований в Лондоне и Терье Лёмо описали совместную работу, проведенную ими в лаборатории Пера Андерсена в Осло. Они обнажали у наркотизированных кроликов гиппокамп и идущие к нему нервные пути и подводили к одному из этих путей - «перфорантному» (perforant) - стимулирующие электроды, а регистрирующие электроды вводили в ту область гиппокампа, где перфорантный путь образует синапсы (в зубчатую извилину, рис. 9.4). Когда они после этого стимулировали перфорантный путь залпом электрических импульсов частотой 10-100 в секунду и длительностью до 10 секунд, наблюдалось необычайно продолжительное (до 10 часов) усиление активности нейронов зубчатой извилины гиппокампа [15]. Авторы назвали этот феномен долговременной потенциацией (сокращенно ДВП). На самом деле ДВП длится гораздо дольше десяти часов; этот эффект наблюдали и у наркотизированных животных с вживленными электродами, причем в этом случае потенциация сохранялась иногда на протяжении 16 недель после первоначальной кратковременной стимуляции. По-видимому, краткая электрическая стимуляция гиппокампа приводила к изменению электрических свойств его клеток.
Блисс, Лемо, Андерсен и многие другие нейробиологи сразу же заинтересовались этим явлением. Это был сильно выраженный эффект - специфический, воспроизводимый и, сверх того, поддававшийся физиологическому, а позднее также биохимическому, фармакологическому и морфологическому исследованию. Гиппокамп млекопитающих был уже хорошо известной структурой, его нервные связи, входные и выходные нервные пути подробно картированы и легко распознаваемы на различных препаратах, хотя индивидуальные нейроны не поддавались такой прямой идентификации, как у аплизии.
Продолжительное изменение выходной клеточной реакции на определенное входное воздействие служит по меньшей мере ярким примером нервной пластичности; больше того, весьма специфичная форма, которую приобретает реакция, может рассматриваться как проявление памяти. Было уже достаточно хорошо известно, что и у человека, и у других млекопитающих гиппокамп имеет какое-то отношение к памяти. Не может ли долговременная потенциация (ДВП) служить механизмом образования энграмм? Не могут ли физиологи использовать ее хотя бы как модель для изучения памяти? Блисс и Лемо явно склонялись к этому, считая свою процедуру стимуляции аналогом «условного раздражения»; однако они заключили свою статью загадочной оговоркой: «Другое дело - использует ли интактное животное в реальной жизни то свойство, которое было выявлено путем синхронной повторной стимуляции группы нервных волокон, нормальная активность которых неизвестна» [15].
ДВП легко вызывать и изучать классическими методами нейрофизиологии, поэтому вряд ли стоит удивляться ее популярности в качестве потенциальной модели памяти. В ближайшие годы после первых наблюдений Блисс в Лондоне, Андерсен в Осло и все большее число исследователей в других лабораториях стали в мельчайших подробностях изучать физиологию ДВП. Было показано, что этот феномен выявляется не только у наркотизированных и ненаркотизированных кроликов, крыс и других лабораторных животных, но и в препаратах in vitro.
Гиппокамп легко извлечь из мозга вместе с его входными нервными путями, в том числе и перфорантным путем. Благодаря трехмерной структуре гиппокампа можно получать тонкие срезы с интактными путями к клеткам того же среза, как показано на рис. 9.4. Такие срезы культивировали in vitro и изучали их электрические свойства. Так забытая с пятидесятых годов методика Мак-Илвейна снова вошла в моду у нейробиологов в конце семидесятых годов. Надлежащая стимуляция нервных путей в таких срезах тоже приводила к ДВП, которая сохранялась до отмирания среза.
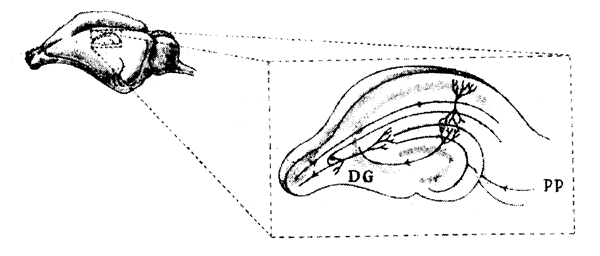
Рис 9.4. Гиппокамп крысы. На увеличенном изображении представлен срез гиппокампа с зубчатой извилиной (DG) и перфорантным нервным путем (РР) Другие области, где имеет место ДВП, включают СА1 и САЗ
Характеристики ДВП примерно одинаковы и в срезах, и в интактном мозгу. Во-первых, в обоих случаях реакция специфична для определенных путей. Иными словами, она развивается только в тех клетках, к которым подводится «условный раздражитель», а не распространяется от клетки к клетке, т. е. это результат функционирования сети специфических связей, а не волна диффузной активности. Такую специфичность можно изящно продемонстрировать благодаря наличию нескольких различных подводящих путей к отдельным участкам даже в одном срезе гиппокампа. Во-вторых, для индукции ДВП требуется повторное воздействие достаточно частых импульсов: то же число импульсов, поступающих с меньшей частотой, не вызывает эффекта. Таким образом, существует частотный порог индукции ДВП. При частоте импульсов выше пороговой ДВП может развиваться либо постепенно, либо по принципу «все или ничего» в зависимости от ритма, интенсивности и частоты условных стимулов. Развитие ДВП происходит по меньшей мере в две (возможно, в три) фазы, с коротким периодом инициации и более длительной фазой устойчивой реакции, которые рассматриваются как аналоги перехода от кратковременной памяти к долговременной.
Третий и, пожалуй, самый интересный момент с точки зрения клеточной аналогии памяти состоит в следующем. В начале восьмидесятых годов было показано, что существует ассоциативная форма ДВП. При этом слабая стимуляция, недостаточная сама по себе для поддержания ДВП, может стать активирующим фактором, если сочетается с сильной стимуляцией через второй путь [16]. Оба вида стимуляции должны совпадать или ассоциироваться во времени, как условный и безусловный раздражители при ассоциативном научении. Можно даже получить такую форму ассоциативного научения, в которой смешаны поведенческие и нейрофизиологические стимулы. Крыса может научиться переходить границу между двумя отделениями ящика в ответ на сигнал, состоящий только из залпа импульсов на гиппокамп, в качестве безусловного раздражителя [17]. Все эти особенности ДВП, взятые вместе, вызывали повышенный интерес к ее изучению по меньшей мере как одной из интригующих моделей памяти.








