 2015-05-26
2015-05-26 2031
2031Эректусы были сравнительно широко распространены по Старому Свету и распадались на ряд локальных подвидов. За африканским подвидом закрепилось название Homo ergaster, хотя к африканским эректусам относят также атлантропа иродезийского человека. За европейским подвидом закрепилось название гейдельбергского человека, хотя существовали и «догейдельбергские» эректусы. В Восточной Азии обитали два подвида: более прогрессивные синантропы из Китая и более примитивныепитекантропы из Индонезии.
Находки на территории России[править | править исходный текст]
Костные остатки эректусов были найдены в том числе и на Русской равнине и Кавказе.[3][4][5] В частности, на Русской равнине, стоянки и орудия эректусов обнаружены вВоронежской[6], Калужской[7], Тульской и Волгоградской областях[8]. Эректусы также были распространены в Восточной[9] и Средней Сибири, в Северо-Минусинской впадине, в Долине Енисея.[10]
До недавнего времени считалось, что эректусы вымерли около 300 тысяч лет назад, уступив место неандертальцам. Однако последние находки свидетельствуют, что они на окраинах ареала могли дожить до прихода современных людей. Последние питекантропы вИндонезии вымерли 27 тыс. лет назад,[11].
Внешний вид[править | править исходный текст]
Эректусы обладали средним ростом (1,5—1,8 м), прямой походкой и архаическим строением черепа (толстые стенки, низкая лобная кость, выступающие надглазничные валики, скошенный подбородок). Объём мозга достигал 900—1200 см³, что больше, чем у Homo habilis, но несколько меньше, чем у Homo sapiens и Homo neanderthalensis.
Материальная культура[править | править исходный текст]
Эректусы активно изготовляли каменные орудия (ашёльская культура), использовали шкуры в качестве одежды, жили в пещерах, пользовались огнём и практиковали каннибализм[12]; около 1,9 миллиона лет назад начали готовить пищу на огне[13].
Находки в Африке, южных районах Европы и Азии (первые находки сделаны на Яве в Триниле Эженом Дюбуа).
Наиболее массивные представители древнейших людей. Являясь уже несомненными людьми, Homo erectus всё же сильно отличались от современного человека, так что многие антропологи склоняются к выделению для них особого рода Питекантропов (Pithecanthropus).
Иногда среди этого типа гоминид выделяют множество видов: Pithecanthropus pekinensis (синантроп) в Китае, Pithecanthropus leakeyi в Восточной Африке, Pithecanthropuspalaeojavanicus на Яве ,Pithecanthropus mauritanicus в Северной Африке и т.д. Видимо, существовало как минимум две основных географических ветви этих гоминид – западная или афро-европейская и восточная или азиатская.
Рядом исследователей Homo erectus объединяется в один вид с более древними Homo ergaster, однако отличия между ними достаточно велики.
От более поздних гоминид Homo erectus отличались в основном заметно более низким черепом и деталями лицевой морфологии.
Эти гоминиды продолжали расселение по планете, достигнув на востоке Индонезии, а на западе - Испании. При этом население, судя по всему, концентрировалось в южных областях и не выходило за пределы субтропического пояса.
Создатели ранней и средней ашельской культуры.
11. НЕАНДЕРТАЛЬЦЫ – ОСНОВНАЯ БИОЛОГИЧЕСКАЯ И КУЛЬТУРНАЯ ХАРАКТЕРИСТИКА. СОВРЕМЕННЫЕ ДАННЫЕ ПО ГЕНЕТИКЕ И АНТРОПОЛОГИИ. ПРОБЛЕМА БИОЛОГИЧЕСКОГО РОДСТВА САПИЕНСОВ И НЕАНДЕРТАЛЬЦЕВ.
Неандерта́лец, человек неандертальский (лат. Homo neanderthalensis или Homo sapiens neanderthalensis; в советской литературе также именовался палеоантропом) — ископаемый вид человека, обитавший 140[1]—24[2] тысячи лет назад. Первые люди с чертами прото-неандертальца, как полагают, существовали в Европе ещё 600 000—350 000 лет назад[3].
История обнаружения[править | править исходный текст]
Первый череп неандертальца был найден в Бельгии ещё в 1829 году. Вторая находка 1848 года связана с английской военной базой на Гибралтаре. Но признаны эти первые находки неандертальскими позднее, в 1836 и 1864 году соответственно[4].
Находка черепа, давшая название новому виду ископаемых людей, является уже третьей по хронологии выявления. Она найдена в 1856 году в Рейнской провинции, недалеко отДюссельдорфа, в Неандерской долине (реки Дюссель, притока Рейна), в небольшом Фельдгоферском гроте, напротив более значительной Неандерской пещеры. Вход в этот грот был скрыт выступом скалы и слоями глины делювиального происхождения, покрывающей здесь всюду известняки. Так как поблизости находились каменоломни, то рабочие дошли до закрывающей грот скалы, взломали её и открыли пещеру, заполненную наносным илом. По удалении этого наноса открылся грот. Недалеко от входа, в глине, рабочие нашли несколько человеческих костей и неполный череп, которые доктор Фульротприобрёл в собственность. Других костей или каких-нибудь изделий в пещерном иле найдено не было, тем не менее нахождение их в древнем наносе, в котором (в других пещерах той же долины, например, в отстоящей всего на 30 шагов так называемой Чёртовой комнате Teufelskammer) были найдены останки несомненно животных делювия, а равно вид костей, степень их сохранности, особенности черепа и т. д. побудили признать в них останки человека глубокой древности, имеющие большое значение для суждения о типе древнейшего европейского населения.
Вирхов признал в этом черепе, а также и в костях конечностей, следы патологических изменений, вызванных рахитом, и высказал мнение, что это не расовый, а индивидуальный тип. Кроме того, он указал на некоторые черепа новейшего времени, выказывающие подобное развитие надбровных дуг и почти столь же покатый взад лоб. С опубликованием эволюционной теории Дарвина в 1859 году антропологи стали рассматривать неандертальцев как «промежуточное звено превращения обезьяны в человека». Современные представления о неандертальских людях отличаются от представлений XIX века.
Особенности физиологии[править | править исходный текст]

«Классический» неандерталец из грота Ля Шапель-о-Сен в реконструкции М. М. Герасимова(Государственный исторический музей, Москва)
Неандертальцы обладали средним ростом (около 165 см), массивным телосложением и большой головой необычной формы. По объёму черепной коробки (1400—1740 см³) они даже превосходили современных людей. Их отличали мощные надбровные дуги, выступающий широкий нос и очень маленький подбородок. Шея короткая и как будто под тяжестью головы наклонена вперёд, руки короткие и лапообразные.[5]Существуют предположения, что они могли быть рыжими и бледнолицыми.[6]
Анализ костного строения показывает, что дети неандертальцев выглядели как маленькие взрослые и, вероятно, их половая зрелость наступала уже к 8-10 годам.[7][8]Средняя продолжительность жизни составляла 22,9 лет.[9] Идентичность гена FOXP2(связанного с речью) у современного человека и неандертальца,[10] а также строение голосового аппарата и мозга неандертальцев, позволяют сделать вывод о том, что они могли обладать речью. По мнению академика В. В. Иванова, у неандертальца «мог быть язык из согласных с малым количеством гласных, что встречается и в человеческих языках»[11].
Неандертальский подросток из пещерыТешик-Таш (Узбекистан) (Герасимов М. М.Биологический музей им. Тимирязева)
Мышечная масса неандертальца была на 30-40 % больше, чем у кроманьонца, скелет тяжелее. Также неандертальцы лучше приспособились к субарктическому климату, поскольку большая носовая полость лучше подогревала холодный воздух, тем самым снижая риск простуды.[12]
Карен Штейдель-Намберс (англ. Karen Steudel-Numbers) из Висконсинского университета в Мадисонеопределила, что вследствие плотного телосложения и укороченной берцовой кости, сокращающей шаг, энергетические затраты на передвижение у неандертальцев были на 32 % выше, чем у современного человека. При помощи модели Эндрю Фрейля (англ. Andrew W. Froehle) из Калифорнийского Университета в Сан-Диего и Стивена Черчилля (англ. Steven E. Churchill) из Университета Дьюка уточнено, что ежедневная потребность в пище неандертальца по сравнению с кроманьонцем, обитавшим в тех же климатических условиях, была больше на 100—350 килокалорий.[13] А специальные химические исследования костной ткани показали, что неандертальцы постоянно ели мясо.[12].
Учёные из Института эволюционной антропологии имени Макса Планка в Лейпциге (Германия) обнаружили у неандертальца ген, препятствующий усвоению молока (лактозы) в зрелом возрасте. Также в ходе исследований выяснилось, что неандертальцам были незнакомы многие наследственные заболевания современных людей — аутизм, болезнь Альцгеймера, синдром Дауна, шизофрения.[14]
Характерные черты и классификация[править | править исходный текст]
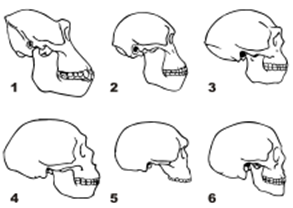
Вид черепа сбоку
1. Горилла 2. Австралопитек 3. Homo erectus 4. Неандерталец (Ля-Шапелль-о-Сен) 5. Штейнгеймский человек 6.Современный человек
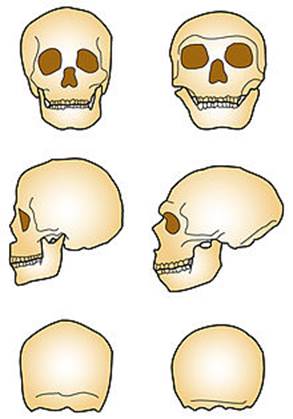
Череп современного (слева) и неандертальского человека (справа)
Неандерталоидность черепа:
· по краниологическим показателям преимущественно долихоцефалия, то есть голова в мозговой части относительно узка и вытянута в длину;
· массивный и утолщённый скелет лицевой и мозговой частей;
· при взгляде сбоку обводы черепа укладываются в окружность с центром в районе уха;
· низкий и как бы убегающий назад лоб без характерных «шишек»;
· мозговая часть невысокая, но зато вытянута назад и вмещает крупный головной мозг;
· очень развитые надбровные дуги;
· широкие скулы;
· лицевая часть массивна, вытянута вперёд (кроме подбородка) и спереди немного заужена;
· при взгляде спереди мозговой части почти не видно, отчего в целом несколько зверообразный вид;
· объёмистая носовая полость и крупная, выдающаяся вперёд переносица, нос может быть с горбинкой;
· длинный скуловой отросток височной кости пропускает мощные жевательные мышцы;
· большая ротовая полость в сочетании с сильными челюстями позволяет откусывать и пережёвывать крупные куски пищи;
· очень крупные челюсти и зубы;
· подбородок почти отсутствует.
Разумеется, эти черты могут встречаться и у современного человека, но обычно по отдельности или реже в том или ином сочетании, причём чаще обращают внимание на большие брови.
Если же мы сравним череп классического неандертальца и не совсем характерный череп нашего современника, имеющего подобные черты (мощные кости, толстые надбровные дуги, глубоко сидящие глазницы, низкий скошенный лоб, мощные челюсти почти без подбородка, покатая и вытянутая назад долихоцефальная форма мозговой части, см. рис. ниже), то можем заметить ещё ряд наиболее характерных отличий:
· в целом на черепе меньше мелкорельефных деталей и формы его, несмотря на грубые черты, более округлы;
· плоское и округлое темя;
· при взгляде с затылка череп округлый (у современного человека — пятиугольный);
· надбровные дуги неандертальца более монолитные и выпуклые, образуют почти сплошной волнообразный валик и вместе с другими обводами глазниц напоминают толстую округлую роговую оправу очков;
· круглые глазные орбиты;
· сглаженные щёчные кости, монолитом соединяющие верхние скулы с верхней челюстью;
· нижняя челюсть не имеет по бокам отростков и при взгляде сбоку вместе с верхней челюстью напоминает круглые кусачки;
· основание черепа имеет очень небольшой изгиб;
· затылок сильно вытянут и заострён наподобие шиньона или горбушки белого хлеба. Снизу на нём развитые выступы, к которым крепились мышцы крепкой и короткой шеи.
Неандертальцы населяли:
· Европу: Неандерталь в Германии, Ла-Шапель-о-Сен во Франции, Киик-Коба в Крыму,Пелопоннес в Греции[17]
· Кавказ: Мезмайская пещера в Краснодарском крае[18]
· Среднюю Азию (Тешик-Таш) и Алтай (пещера Окладникова)[19]
· Ближний и Средний Восток: Кармель в Израиле, Шанидар в иракском Курдистане.
Жилища[править | править исходный текст]
Паола Вилья относит к неандертальскому периоду хижину с костровищем, обнаруженную на территории Ниццы (Терра-Амата), тогда как первооткрыватель памятника Анри де Люмле считает, что хижину построил более ранний гейдельбергский человек.
Согласно исследованиям пещер Бальци-Росси в Италии, проведённым Жюльеном Риель-Сальваторе из Колорадского университета и его коллегами, жилище неандертальцев делилось на три «зоны». В первой из них, в верхней части жилища, располагалась «кухня», о чём свидетельствуют найденные в этой зоне останки животных, следы охры, которая применялась для выделки шкур, в качестве клея и как антисептическое средство. Во второй зоне, которая была самой просторной, размещался очаг и здесь было спальное место. В третьей зоне была «мастерская» где изготавливались каменные инструменты, так как здесь было больше всего солнечного света и минимальный риск, что кто-то помешает работе.[20]
Обычаи[править | править исходный текст]
Согласно анализу митохондриальной ДНК останков двенадцати погибших под обвалом неандертальцев из пещеры Эль-Сидрон на севере Пиренейского полуострова (совр. Испания) выяснилось, что у троих мужчин, одной женщины, двоих подростков и ребёнка 8-9 лет был общий митохондриальный гаплотип, условно обозначенный исследователями как «А» (что говорит о близком родстве). Две остальные найденные взрослые женщины имели митохондриальные гаплотипы, условно обозначенные как «В» и «С», отличные от основного в группе. Также общий с одной из этих женщин гаплотип «С» был у подростка и детей возраста 2-3 и 5-6 лет. Этот анализ свидетельствует о патрилокальности неандертальцев, то есть о том, что юноши, достигшие половой зрелости, оставались в группе, а девушки — переходили в семью мужа, а также о близкородственных браках внутри группы. Минимальный интервал между родами у неандертальских женщин (судя по возрасту детей с гаплотипом «С») сопоставим с таковым в племенах современных охотников-собирателей, у которых он в среднем составляет 3,4 года.[21]
Искусство[править | править исходный текст]
Неандертальцам приписывается самый ранний известный музыкальный инструмент — костяная флейта с 4 отверстиями (англ.). Однако проведённый анализ краёв отверстий показал, что отверстия могут быть и следами зубов гиены,[22] поэтому «флейта» могла иметь и естественное происхождение.
Неандертальцы умели использовать самодельные инструменты и оружие, но, по-видимому, у них не было никакого метательного оружия[23] — добычу убивали ударом коротких копий, о чём свидетельствуют следы развитых мышц на костях правой руки.
В 2003 году на берегу Луары в местечке Ля Роше-Котар (фр. La Roche-Cotard) был найден камень приблизительно 10 см высотой, имеющий поразительное сходство с человеческим лицом. Камень датирован 35 тыс. лет назад. Эта важная находка является свидетельством, что у неандертальцев было представление об искусстве.[24] Впрочем, аналогичных «личин» на других стоянках пока не обнаружено, что не позволяет сделать однозначный вывод о существовании подобного рода искусства.[25]
С неандертальцами также связаны параллельные царапинки на костях (Арси-сюр-Кюр, Бачокиро, Молодова), ямки на каменной плите (Ля Феррасси) неясного назначения. Достаточно редки находки украшений в виде просверленных зубов животных, датируемых около 30-34 тыс. лет назад. А единственный образец настоящего рисунка представляет собой процарапанное на кости изображение леопарда, найденное на стоянке Пронятин (Украина) с датировкой около 30-40 тыс. лет назад.[22] Обнаруживаются также принадлежащие неандертальцам продырявленные и раскрашенные ракушки, служившие, по-видимому, украшением.[25]
В ходе раскопок стоянки неандертальцев возле Вероны палеонтолог Марко Перезани (Marco Peresani) из университета Феррары(итал. Università degli Studi di Ferrara) и его коллеги обнаружили останки 22 видов птиц, у которых были срезаны перья. Идентифицированыбородач-ягнятник, кобчик, чёрный евразийский стервятник, золотой орёл, лесной голубь и альпийская галка. Окрас этих птиц варьируется от чёрного и серого до голубого и оранжевого, а перья достаточно длинные. Предполагается что неандертальцы украшали себя перьями, варьируя цветовой орнамент. Результаты исследования были опубликованы в журнале Proceedings of the National Academy of Sciences (PNAS).[25][26]
Религия[править | править исходный текст]
Неандертальцы являлись носителями мустьерской культуры. В гроте Ля-Шапель-о-Сен во Франции было обнаружено неглубокое захоронение со скелетом в позе эмбриона. Рядом с телом были оставлены цветы, яйца и мясо, что может свидетельствовать о вере взагробную жизнь и наличии религиозно-магической практики.
Некоторые погребальные обычаи неандертальцев и современных людей близки. К примеру, в Ираке было обнаружено погребение 40-летнего мужчины, на которое соплеменники возложили цветы — вокруг скелета находилось много цветочной пыльцы. Другие обычаи разнятся. Так, неандертальцы, жившие в суровых условиях ледникового периода, часто съедали своих умерших сородичей — из 300 найденных скелетов хорошо сохранились только 12. Остальные имеют следы отделения мяса, в костях содержатся крошечные осколки кремнёвых орудий[12].
Религиозная практика европейских неандертальцев представлена также «культом медвежьих черепов», что позволяет предполагать существование обрядов охотничьей магии. На стоянке Ильская существовал культ черепов зубра.[22]
Культура неандертальцев (т. н. «мустьерская», или, что то же самое, среднепалеолитическая культура) — это прежде всего обоюдоострые рубила, заточенные более качественно, чем аналогичные орудия H.erectus, а также разнообразные отщепы, использовавшиеся для разделки туш. У неандертальцев имелись также деревянные копья с каменными наконечниками для ближнего боя. В более позднее время, уже во время контактов с сапиенсами (см. ниже), у неандертальцев появляются зачатки искусства (ожерелье из медвежьих когтей, нечто вроде «флейт» — кости с просверлёнными дырочками, которые, впрочем, могли служить для разведения огня, а не для музыкальных упражнений.[27]
Наука (медицина)[править | править исходный текст]
Во всех человеческих цивилизациях есть аналог профессии костоправа — человека, который профессионально занимается восстановлением сломанных конечностей людей и животных, а также восстанавливает суставы при вывихах. Так, при анализе 36 скелетов неандертальцев, имеющих переломы, только у 11 результаты лечения перелома признаны неудовлетворительными. Это показывает, что уже на таком уровне развития эффективность медицинской помощи при переломах превышала 70 %, первобытные люди знали о переломах и умели их лечить.[28]
Находка в пещере Шанидар (Ирак), датируемая 50 тысячами лет, показала, что неандертальцы заботились о раненых соплеменниках. При обвале в гроте погибли двое калек, которые, очевидно, находились там одни, пока остальные члены группы охотились или занимались другими делами. У одного из них были свежие раны на рёбрах, у другого заживающий перелом черепа.[12]
Родство с современным человеком[править | править исходный текст]

Неандертальцы у костра под открытым небом (компьютерная графика)
Современные учёные больше не считают неандертальцев предками современных людей[29], поскольку, согласно данным генетики, прямые предки людей происходят изАфрики, которая лежала вне ареала неандертальцев (см. статью Митохондриальная Ева). В 1997 году на основании анализа ДНК первого неандертальца учёные Мюнхенского университета сделали вывод: различия в генах слишком велики, чтобы считать неандертальцев предками кроманьонцев (непосредственных предков современных людей). Эти выводы были подтверждены ведущими специалистами из Цюриха, а позже и другими специалистами из Европы и Америки.[30]
В 2006 году началась расшифровка генома неандертальцев.[31] Выяснилось, что генетическое расхождение между современными людьми и неандертальцами произошло около 500 тысяч лет назад, то есть ещё до распространения ныне существующих рас человека.[32]
Тем не менее, два вида людей сосуществовали многие тысячи лет.[33] В частности, на стоянках и неандертальцев, и кроманьонцев были обнаружены обглоданные кости другого вида.[34] Профессор Бордосского Университета Жан-Жак Юблен считает, что два вида людей преимущественно враждовали.
Однако есть также доказательства смешения между неандертальцами и современными людьми.[35] В 2009 году профессор Сванте Паабо изИнститута эволюционной антропологии Макса Планка в Лейпциге сообщил на ежегодном собрании Американской ассоциации по развитию науки об успешном прочтении ядерного генома неандертальца. Первоначально каких-либо признаков гибридизации кроманьонцев с неандертальцами не удалось обнаружить.[36] Однако уже к маю 2010 года гены неандертальца были найдены в геномах целого ряда современных народов.[37][38] «Те из нас, кто живёт за пределами Африки, несут некоторое количество ДНК неандертальца», — заявил профессор Паабо. «Генетический материал, унаследованный от неандертальцев, составляет от 1 до 4 %. Это немного, но достаточно, чтобы утверждать о достоверном наследовании существенной части признаков у всех из нас, кроме африканцев» — сообщил доктор Давид Райх из Гарварда, также участвовавший в работе. В исследовании геном неандертальца сравнивали с геномами пятерых наших современников из Китая, Франции, Африки и Папуа — Новой Гвинеи. Скрещивание было зафиксировано и, вероятно, произошло вскоре после миграции предков современного человека из Африки, то есть на территории Ближнего Востока, поскольку у трёх человек из разных регионов мира, за исключением Африки, пропорция генов неандертальца примерно одинакова. Обратный дрейф генов, от современного человека к неандертальцам, обнаружен не был. Это объясняется взаимодействием небольшой группы африканских колонистов с многочисленной популяцией неандертальских аборигенов, населявших в то время Ближний Восток. В то же время, авторы исследования не могут полностью отбросить и альтернативное объяснение присутствия генов неандертальцев у современных людей, согласно которому небольшая группа, от которой произошло все население Евразии, Америки и Австралии, изначально генетически отличалась от остального африканского населения и была ближе к неандертальцам, чем остальные. Скрещивание является лишь наиболее вероятной версией.[37] Недавно палеонтологам удалось найти на территории северной Италии останки гибрида неандертальца и Homo sapiens — единственного известного на сегодняшний момент. Гоминид жил приблизительно 30-40 тысяч лет назад[39].
Геном[править | править исходный текст]
Основная статья: Расшифровка генома неандертальца
Геном неандертальца по размеру близок к геному современного человека. Предварительные результаты показывают, что ДНК современного человека и неандертальца идентичны приблизительно на 99,5 %. Исследователи извлекли ископаемую ДНК неандертальца из кости бедра скелета 38000-летней давности мужчины-неандертальца из пещеры Виндия в Хорватии, а также из других костей, найденных в Испании, России и Германии.[40][41]
12. ПОНЯТИЕ «РАСА» В АНТРОПОЛОГИИ. ХАРАКТЕРИСТИКА БОЛЬШОЙ ЕВРОПЕОИДНОЙ РАСЫ.
Путешественники, исследовавшие разные области Земного шара, всегда неизбежно сталкивались с пестротой населяющих его народов. Различия людей разных регионов часто выглядят весьма значительными. Незнакомым и необычным часто кажется буквально все - одежда, язык, хозяйство, поведение и, наконец, внешность - особенности морфологии и физического строения, подлежащие исследованию биологической антропологией. Однако среди фейерверка цветов, форм и звуков можно заметить определенную систему. В ходе длительных дискуссий и горячих споров в науке оформилось представление о разных способах классификации групп человека. Впрочем, и сейчас единые законы такого деления признаны далеко не всеми, и сами его основы подвергаются значительным сомнениям. Один из аспектов биологической изменчивости географических групп современного человека изучает расоведение. Соответственно, географические группы людей, выделяемые по биологическим признакам, называются расами. Под термином "раса" понимается система человеческих популяций, характеризующихся сходством по комплексу определенных наследственных биологических признаков (расовых признаков). Важно подчеркнуть, что в процессе своего возникновения эти популяции связаны с определенным географическим ареалом и естественной средой. Это конечно, не единственное из возможных определений - суть понятия можно уточнять, описывать при помощи терминов фенетики и генетики.
Например, можно сказать иначе: раса - это система популяций, обладающих достаточным фенотипическим и генетическим сходством, позволяющим отличить их от других подобных систем; расы - открытые генетические системы, в результате обмена генами между которыми могут возникать смешанные популяции.
Суть понятия, во всех этих определениях остается неизменной - термин определяет единицу внутривидовой, и при этом надпопуляционной структуры современного человечества, выделенную по определенному набору (комплексу) наследственно обусловленных признаков и занимающую определенный ареал. С точки зрения основ современной систематики, расы соответствуют уровню подвидов, несомненно, принадлежащих к одному единому биологическому виду Homo sapiens. Отличие же рас от подвидов животных, связаны с общественной природой человека. Наконец, раса принципиально отличается от этноса биологическими критериями выделения, хотя в силу исторических причин иногда наблюдается частичное совпадение расовых групп и конкретных этносов. Мы поговорим об этом ниже, пока же важно запомнить, что раса понятие сугубо биологическое, как и сами признаки, по которым проводится расовая классификация.
Понятия расы, а также различные пути группирования людей, изменяются культурой и временем, на них также влияют общественные и политические причины.
Раса – это признак унаследованный (то есть генетический), на формирование которого повлияли общественные и культурные показатели
ЕВРОПЕОИДНАЯ, или европейско-азиатская, большая раса, которую в живой речи нередко называют «белой», чрезвычайно многочисленна и охватывает около 40% человечества, т. е. около 1,15 млрд. людей. С открытием Америки, а позже и Австралии европеоиды расселились по всему слету. Однако главное ядро расы находится в пределах Старого Света —в Европе, Азии и Сонорной Африке. В одной лишь Индии насчитываются многие десятки миллионов индийцев и других европеоидов. Приведем общую антропологическую характеристику европеоидной расы. Окраска кожи от светлого до смуглого, даже коричневого цвета; волосы на голове мягкие, волнистые (или прямые), варьируют по цвету от светлых до темных, третичный волосяной покров на теле развит сильно или средне, на лице (борода и усы) нередко сильно; лоб прямой или несколько наклонный. Лицо в среднем отделе довольно сильно выступает вперед, но скулы мало выдаются, равно как и челюсти; и общем лицо ортогнатное; углы глаз располагаются на одном уровне, складка верхнего века развита слабо; у большинства глаза карие, встречаются и светлые —серые, синие и голубые, обычные у народов северной половины Европы; нос узкий, обычно сильно выступающий, с довольно высоким переносьем; продольные оси наружных носовых отверстий обращены почти прямо вперед (положение ноздрей сагиттальное); губы тонкие или средней толщины, не выступают вперед (ортохейлия); подбородочный выступ развит средне или сильно; форма головы очень изменчива; широко распространены как брахикефальные или мезокефальные, так и долихокефальные типы. В большую европеоидную расу входят две малые расы: южная европеоидная, или индо-средиземноморская, и северная европеоидная, или атланто-балтийская. Первая имеет более темно-,а вторая —более светлоокрашенные кожу, волосы и радужину глаз. Обе расы соединяются между собой переходными группами антропологических типов, которые, характеризуясь темными волосами, брахицефалией, средним ростом, образуют, по Я. Я. Рогинскому (1956), среднеевропейскую расу. В качестве представителей индо-средиземноморской расы можно назвать индийцев, таджиков, армян, греков, арабов, итальянцев, испанцев. Для них характерны: черные, волнистые волосы, карие глаза, нос с выпуклой спинкой; лицо очень узкое, форма головы долихо- или мезокефальная. У русских, белорусов, поляков, норвежцев, немцев, англичан и других европейских народов, живущих более к северу, часто встречается иная совокупность черт: очень светлая кожа, белокурые или светлорусые волосы; серые или голубые глаза, относительно длинный нос. Эти высокорослые европейцы входят в состав атланто-балтийской расы.
13. АДАПТИВНЫЕ ЧЕРТЫ В ХАРАКТЕРИСТИКЕ ЧЕЛОВЕЧЕСКИХ РАС. ХАРАКТЕРИСТИКА БОЛЬШОЙ МОНГОЛОИДНОЙ РАСЫ.
Легко заметить, что в странах с жарким климатом цвет кожи людей заметно темнее, чем в странах с холодным. Также, ближе к экватору увеличивается ширина носа, толщина губ, выступание лица вперед. Такое распространение признаков объясняется следующим образом.
В глубокой, древности, вероятно, в верхнем палеолите, биологическое приспособление к условиям окружающей среды имело очень большое значение. Человек был еще не настолько вооружен технически, чтобы эффективно оградить себя от действия стихий. В таких условиях цвет кожи, например, играл жизненно важную роль для теплообмена. Темный пигмент кожи поглощает значительно большее количество ультрафиолета, предохраняя организм от опасности раковых заболеваний. Хотя темная кожа сильнее нагревается от солнечных лучей, более эффективная работа кожных желез спасает от перегрева. Температура кожи негроида в одинаковых условиях меньше, чем у европейца или азиата. Светлая кожа более подвержена ультрафиолетовому излучению. Это может быть полезным в северном полушарии, поскольку в условиях сравнительно малой инсоляции – солнечной освещенности – возникает проблема рахита – недостатка D. Т.о., в северных широтах увеличение чувствительности кожи к ультрафиолету спасает детей от рахита. Приспособительное значение имеют, курчавые волосы и вытянутая высокая форма головы экваториальных групп населения. Курчавые волосы создают на голове воздушную подушку, а воздух – один из лучших теплоизоляторов. Кроме того, в волосе негра содержится больше число пузырьков воздуха, чем в волосе монголоида или европеоида. Вытянутая узкая и высокая голова экваториальных групп имеет меньшую верхнюю площадь при том же объеме, что и низкая широкая голова северных народов. Это спасает ее от чрезмерного перегрева на солнце.
Большая ширина рта и значительная ширина слизистой губ негроидов полезны в жарком климате, поскольку увеличивают поверхность испарения влаги, охлаждая организм. Уплощенный широкий нос имеет аналогичное значение. Вместе с тем, малые размеры носовой полости не позволяют воздуху дополнительно нагреваться при вдохе.
Противоположное значение имеет высокий длинный нос европеоидов и северных монголоидов. Проходящий через длинный носовой ход воздух успевает нагреться и попадает в легкие теплым. Ортогнатизм северных рас – укороченность лица –увеличивает изгиб носового хода и предохраняет носоглотку от переохлаждения.
Ширина глазной щели заметно отличается у представителей разных рас. Узкий разрез глаз монголоидов, бушменов и туарегов предохраняет глазное яблоко от пыли, ветра и слишком яркого солнца на открытых пространствах. Вместе с этим, глаз монголоидов существенно отличается большим количеством подкожной клетчатки, предназначенной для сохранения тепла.
Более экзотическим примером адаптивного значения расовых признаков может служить стеатопигия бушменов, готтентотов и андаманцев. Жировые отложения на ягодицах и бедрах служат запасом питательных веществ в условиях их резкого недостатка. Благодаря скульптурам верхнего палеолита мы знаем, что стеатопигия была обычным явлением в Европе около 25-20 тыс. лет назад, во время становления расовых комплексов
Классические расовые признаки включают черты внешности – цвет и форму глаз, губ, носа, волос, цвет кожи, строение лица в целом, форму головы.
МОНГОЛОИДНАЯ, или азиатско-американская, большая раса, которую иногда по устарелой терминологии называют «желтой», охватывает около 50% всего населения земного шара, то есть 1,3 млрд. Из этого количества почти половину составляют китайцы —около 600 млн. человек. Главная масса представителей «желтой» расы занимает огромные пространства Азии, в особенности ее северные, восточные, центральные и юго-восточные области. Распространилась монголоидная раса также в Океанию и на материк Америки. Очень многие монголоидные группы входят в состав населения СССР, преимущественно его Азиатской части, где характерными представителями их являются якуты, буряты, тунгусы (эвенки), чукчи, тувинцы, алтайцы, гиляки (нивхи), алеуты, азиатские эскимосы и многие другие. В Европейской части СССР монголоидные антропологические типы входят в состав башкир, татар, чувашей и некоторых других народов. Для большой монголоидной расы характерны следующие признаки: кожа светлая или смуглая, с желтоватым или желтовато-коричневым оттенком; волосы на голове почти у всех прямые и жесткие (тугие), обычно черного цвета; борода и усы, как правило, развиваются поздно а слабо; волосы третичного покрова на теле почти отсутствуют. У представителей многих антропологических типов этой расы, особенно у северных монголоидов, лицо крупное, вперед выступает средне (мезогнатизм); в связи с разрастанием и выступанием скул в стороны оно значительно уплощено; глаза карие, их разрез у большинства средний, но у многих узкий, причем наружный угол глазной щели располагается нередко выше внутреннего; складка верхнего века развита сильно, у очень многих почти достигает ресниц, переходит на нижнее веко и полностью или частично прикрывает внутренний угол глаза, захватывая слезный бугорок: тут образуется особая складка —эпикантус; нос средней ширины, выступает слабо, обычно с низким переносьем (у индейцев выступает сильно, переносье высокое, у эскимосов —очень низкое); положение ноздрей у большинства среднее; губы тонкие или средней толщины, верхняя прохейлична; подбородочный выступ развит средне; голова у очень многих мезокефальная. Монголоидная большая раса делится на три малые расы. Первая из них —северная монголоидная, или азиатская континентальная; вторая —южная монголоидная, или азиатско-тихоокеанская; третья —американская (индейская). Представителями северной монголоидной, или, как ее еще называют, центрально-азиатской, малой расы являются, например, буряты и монголы. Это довольно типичные монголоиды, которые, однако, отличаются несколько ослабленными чертами, так как у них цвет кожи, волос и глаз более светлый, волосы не всегда тугие; но борода почти не вырастает, губы тонкие, лицо большое и плоское. На юго-востоке Азии преобладает южная монголоидная раса, у большинства представителей которой —малайцев, яванцев, зондцев —кожа более смуглая; лицо более узкое и низкое; губы средней толщины идя толстые; нос широкий; эпикантус встречается реже, чем у северных монголоидов; борода развивается, хотя и не сильно; па голове у некоторых волнистые волосы; рост несколько ниже, чем у северных монголоидов, и значительно ниже, чем у китайцев. Третья монголоидная раса —американская (индейская) —обнаруживает переходный характер, так как обладает более слабо выраженными монголоидными чертами и в то же время некоторыми особенностями, сближающими се с европеоидным типом. У индейцев волосы обычно прямые и тугие, черного цвета; борода, усы, третичный волосяной покров тела развиваются слабо; кожа желтовато-коричневая, глаза темные, карие; лицо у очень многих широкое. По этим признакам индейцы сходны с типичными монголоидами. Однако по складке верхнего века (которая хотя и развита сильно, но эпикантуса, как правило, не образует), по сильно выступающему носу, часто с выпуклой спинкой, по средней или большой высоте переносья, а также по общему типу лица индейцы напоминают европеоидов. У некоторых племен отмечают волнистость волос на голове
14. ТИПОЛОГИЧЕСКАЯ И ПОПУЛЯЦИОННАЯ КОНЦЕПЦИИ РАСОВЕДЕНИЯ. ХАРАКТЕРИСТИКА БОЛЬШОЙ НЕГРОИДНОЙ РАСЫ.
Типологическая и популяционная концепции расоведения. Характеристика большой негроидной расы.
Типологическая концепция расоведения.
Типологическая концепция рас, признающая гипотезу наследования расовых признаков, откуда делается вывод о совпадение морфологических особенностей индивидуума с расовой категорией к которой он принадлежит. Согласно типологического подходу описав черты конкретного человека, можно четко отнести его к той или иной расе. Выделяются расовые типы, и каждый индивид оценивается по степени приближения к тому или иному «чистому» типу. По такой системе можно определять расовую принадлежность человека в процентах.
Сложность данной концепции заключается в выделение «чистых» типов. В рамках типологической концепции смешивается расовая изменчивость и изменчивость индивидуальная. Близкие родственники могут быть описаны как принадлежащие к разным расам, и наоборот люди весьма далекие географически и родственно относятся к одной группе.
«Другой существенный недостаток индивидуально- типологической методики заключается ы том, что она полностью пренебрегает широко известным и твердо установленным фактом эволюционной биологии – изменчивостью, проявляющейся в любой, даже гомогенной популяции» - Алексеев, 1969год.
«В морфологическом типе индивидуума проявляются физиологически скорелированные связи меду признаками, в то время как исторические закономерности проявляются именно на групповом или популяционном уровне. Поэтому типологический подход к исследованию расовых особенностей популяции всегда связан с риском принять проявления индивидуальной изменчивости за групповую.» - Алексеева, Яблонский, 2002 год.
Популяционная концепция расоведения.
Согласно популяционной концепции, раса – это совокупность популяций, а не индивидуумов. Отнесение конкретного человека к тои или иной рае определяется не столько его индивидуальными чертами, сколько фактом принадлежности к той или иной популяции. Расовые признаки не наследуются единым комплексом, а у индивидуума они совершенно не обязательно соответствуют какому-то характерному типу.
Оценивая изменчивость внутри популяции или группы популяций, можно определить некий образ, который можно назвать расой. Границы рас из-за индивидуальной и популяционной изменчивости очень размыты, между расами так же имеются переходы.
В «соседских» популяциях часто встречаются очень похожие друг на друга индивиды. Однако исследуя популяции в целом, между ними будут обнаружены неуловимые различия. Именно эти значительные сходства некоторых групп популяций и их отличие от других, дают возможность выделять их в расы.
Характеристика большой негроидной расы.
Негроидная раса характеризуется темной окраской кожи, богатой меланином. От коричневого до темно-коричневого.
Рост тела имеет большой разброс, к этой расе относятся представители самых высоких и самых низких племен.
Конечности удлинённые, в частности руки.
Голова долихо-мезокефальная, иногда умеренно брахикефальная (см. в конце билета пояснение.), с прямым, высоким, выпуклым и выступающим лбом и слабо развитым надбровьем. Надбровные и глазничные дуги не выделяются.
Волосы у представителей этой расы темные, густые курчавые и волнистые, не только на голове но и на теле. Оволосение лица у мужчин слабо выражено.
Лицо широкое, прогнантное на счет выступания челюстей, зубов и носовых костей. В целом лицо имеет среднюю или сильную горизонтальную профилировку (степень выступания лица и его частей в вертикальной или горизонтальной плоскостях). Нижняя челюсть лишена подбородочного выступа.
Нос широкий, маловыступающий со средневысокой или низкой переносицей. Ноздри расширены в поперечном направлении.
Губы большие, толстые с большой слизистой частью.
Глаза крупные, в основном карие. Склеры имеют желтоватый оттенок.
Головной индекс.
Отношение максимальной ширины черепа к максимальной длине. Из этого следует деление на:
· Брахикефалия (короткоголовость) — соотношение длины и ширины головы, при котором ширина составляет более 80,9 % длины. К брахицефалам традиционно относят балтидов, альпинидов, арменоидов и монголоидов.
· Мезокефалия (среднеголовость) — градация черепного указателя (76—80,9 %), характеризующая умеренно длинный и широкий череп.
· Долихокефалия (длинноголовость) — форма головы, при которой отношение максимальной ширины головы к максимальной длине (головной указатель) составляет 75,9 % и ниже. Соответствует долихокрании при измерении этих размеров на черепе человека (черепной указатель 74,9 % и ниже). К долихоцефалам традиционно относят представителей нордической и средиземноморской расы.
15. РАСЫ И НАРОДЫ. ОЦЕНКА РАЗЛИЧИЙ В ПОНЯТИЯХ ТЕРМИНОВ. ОЧАГИ И ФАКТОРЫ РАСООБРАЗОВАНИЯ.
Раса – это верхний предел родства популяций людей, высшее общечеловеческое родство в рамках одного вида. Это группа родственных популяций людей, для которых одновременно выполняются 4 условия:
1. Все популяции ограничены родственностью физического типа.
2. Общность физических признаков базируется на законах наследственности.
3. Для всех популяций характерна общность биологической истории.
4. Для каждой расы характерен определенный географический ареал.
Расы отличаются друг от друга физическими признаками – цветом кожи, волос, глаз, формой волос, чертами лица, ростом, формой черепа и др. Физические особенности, по которым одна раса отличается от другой, наследственны и сравнительно мало изменяются в результате непосредственного влияния среды.
Расовые варианты, в отличие от всех других внутривидовых вариантов, приурочены каждый к определенной территории или ареалу данной расы; в настоящее время связь с ареалом иногда не обнаруживается с полной отчетливостью, однако она всегда имела место в более или менее далеком прошлом.
Народ.
Под народом понимают, в частности, этнос, то есть определённую группу людей, отличающуюся общностью ряда признаков — языка, культуры, территории, религии, исторического прошлого и т. Д
Расы и этнос
Большие расы человека занимают обширные территории, охватывающие народы, которые различаются по уровню экономического развития, культуре, языку. Нет сколько-нибудь четких совпадений понятий «раса» и «этнос» (народ, нация, национальность). В то же время есть примеры антропологических типов (малые, а иногда и большие расы), которые соответствуют одному или нескольким близким этносам, например, лапаноидная раса и саамы. Гораздо чаще, однако, наблюдается обратное: один антропологический тип широко распространен среди многих этносов, как, например, в коренном населении Америки или у народов Северной Европы. Вообще все крупные народы, как правило, неоднородны в антропологическом отношении. Нет также совпадения между расами и языковыми группами — последние возникали позже, чем расы. Так, среди тюркоязычных народов имеются представители как европеоидов (азербайджанцы), так и монголоидов (якуты). Термин «расы» неприменим к языковым семьям — например, нужно говорить не о «славянской расе», а о группе родственных народов, говорящих на славянских языках.
Факторы расообразования, это:
-приспособление (адаптация);
-изоляция;
-метисация;
-автогенетические процессы.
Как возникли расы: изоляция
Всего несколько тысяч лет назад население Земли, а соответственно, и плотность этого населения, были невелики. При этом освоенная людьми площадь Земли мало отличалась от современной. Люди еще во времена верхнего палеолита, добрались до всех пригодных к жизни уголков планеты. Отдельные группы людей при этом надолго теряли всякие контакты с остальным человечеством. Такое существование без связей с внешним миром называется изоляцией. У людей она может быть обусловлена двумя разными причинами - географической недоступностью области (географическая изоляция) или социальными факторами (культурная изоляция).
Обычно изоляция групп людей происходила в каких-то особо труднодоступных областях - на удаленных островах, в горных долинах, непроходимых лесах. При этом изоляция могла быть разных масштабов. Например, индейцы обеих Америк тысячи лет были практически полностью изолированы от людей Старого Света, однако на территории целых двух континентов! Вместе с тем, небольшие группы в Андах и амазонской сельве были, в свою очередь, полностью изолированы от других индейцев. Аборигены Австралии также не имели связей с людьми остального мира много тысяч лет, но внутри континента не существует непреодолимых географических барьеров, даже знаменитые австралийские пустыни можно обойти вокруг. Но и в Австралии есть группы, почти полностью изолированные от остального аборигенного населения, например, племена пигмеев барринес в горах восточного побережья.
Иногда географический барьер имеет несколько иной характер. Таковы условия изоляции пигмеев в Африке. Между тропическими дождевыми лесами, в которых живут пигмеи, и саваннами, где живут их соседи банту, нет какой-либо непреодолимой границы. Этой границей служит огромная разница условий жизни в лесу и саванне. Высокорослые люди не могут жить в дождевом лесу, а низкорослым очень трудно выжить в саванне. Культурный и торговый обмен при этом происходит регулярно, однако обмен генами между жителями лесов и саванн довольно слаб, тем более что он направлен преимущественно от пигмеев к банту. Хотя в отдельных местах и возникают смешанные популяции, характерные типы пигмеев и жителей саванн не теряют своей специфики.
Совсем другая ситуация существует в случаях т.н. культурной изоляции. Культурная изоляция возникает, когда в обществе существуют некие запреты на общение с людьми из иных групп. Обычай заключать браки исключительно внутри своей группы называется эндогамией. Запреты бывают самые разнообразные, но чаще всего встречаются ограничения, связанные с религией. Менее строги обычно запреты, связанные с разницей социальных групп. Наиболее известным примером такого рода является кастовая система в Индии. Благодаря ее существованию, антропологические отличия социальных групп в Индии весьма значительны и могут достигать расовых масштабов. Благодаря социальной изоляции, в Южной Индии до сих пор существуют антропологически не изменившиеся потомки африканских рабов, завезенных сюда сотни лет назад. Другим примером такого рода могут служить группы староверов, живущие в разных уголках Сибири. Существующие в окружении аборигенного монголоидного населения, эти группы, тем не менее, полностью сохранили свои антропологические особенности. Абсолютно та же ситуация сложилась и с молоканами - представителями религиозной секты, переселенными в Армению в XIX в. Религиозная изоляция привела к полному сохранению всех исходных расовых признаков у молокан.
Действие изоляции может быть разным. Благодаря ей признаки могут долгое время сохраняться неизменными, как, например, массивность черепа и крупные размеры зубов у австралийских аборигенов, или заметно эволюционировать, как, например, появление светлых волос у тех же самых аборигенов. В очень большой популяции или в общности связанных популяций новый признак имеет меньше шансов сохраниться и закрепиться, чем в малой. С другой стороны, популяции, слабо ограниченные в своем распространении, могут сталкиваться с новыми условиями среды и смешиваться с соседними группами, приобретая тем самым новые признаки. В группе же, обитающей долгое время в неизменных условиях среды, нет закономерных причин изменения признаков, и изменения эти могут носить лишь случайный характер. Таким образом, изоляция приводит как к сохранению разнообразия расовых типов, так и к его возобновлению.
Увеличение численности населения Земного шара постепенно привело к тому, что в настоящее время полностью изолированных групп на планете почти не осталось. Однако жители удаленных областей, например, индейцы лесов Южной Америки и аборигены горных районов Филиппин, имеют крайне малые шансы встретиться. Сохраняет свое значение даже в современных крупных мегаполисах изоляция, связанная с этническим и социальным фактором (та же эндогамия). Поэтому изоляционные механизмы продолжают действовать и в современном мире.
Как возникли расы: метисация
Обратное явление представляет метисация (или метизация). Важнейшее ее значение заключается в том, что метисация поддерживает видовое единство человечества. В истории не известно случаев, чтобы длительная изоляция какой-либо группы населения приводила к появлению нового вида человека.
Как известно, важнейшим показателем видового единства - гибридологическим критерием вида - является возможность получения плодовитого потомства. Вся история человечества является подтверждением возможности получения плодовитого потомства между представителями самых разных рас. Смешанные межрасовые группы населяют в настоящее время почти всю Сибирь, в качестве наиболее ярких примеров можно вспомнить целые народы долганов и камчадалов, образовавшиеся в результате смешения русских поселенцев и аборигенных народов - эвенов и ительменов. В Южной Африке существует народность реоботийских бастардов, представляющих потомков голландских колонистов и аборигенов - готтентотов.
Более того, даже некоторые расы представляют собой результат смешения других рас. Таковы уральская раса, южно-сибирская раса, эфиопская раса, дравидийская раса.
Целые страны в Центральной и Южной Америке и других частях мира населены народами, образовавшимися в ходе смешения черт сразу трех больших рас - монголоидной, европеоидной и негроидной. Достоверная классификация этих групп еще не проведена, хотя такие попытки антропологами уже осуществлялись. Можно говорить о возникновении сразу нескольких смешанных рас буквально за последние несколько столетий. И еще несколько рас можно сейчас наблюдать в процессе их становления!
Метисация приводит к двум противоположным явлениям:
с одной стороны, смешение разных антропологических типов приводит к сглаживанию их различий;
с другой стороны, образующийся смешанный (метисный) вариант, как правило, оказывается отличающимся от исходных.
Признаки в смешанной группе не представляют собой среднее между состояниями признаков в первоначальных группах. Тем более, не усредняется весь комплекс признаков. Таким образом, образуется новый вариант, обособляющийся в новый антропологический тип. Часто такой новый вариант попадает в условия относительной или полной изоляции, где новая комбинация признаков развивается и закрепляется.
При исследовании конкретных групп населения бывает трудно отличить результат метисации и автохтонного развития. Более того, эти процессы часто идут параллельно. Так, ученые не могут прийти к единому мнению по поводу путей происхождения уральской и эфиопской рас. Согласно одной точке зрения, эти расы сложились на той территории, где распространены сейчас, в ходе независимого длительного развития, а их морфологические комплексы своеобразные и весьма древние. Согласно же другому мнению, и уральская, и эфиопская расы представляют собой продукт смешения антропологических типов соседних территорий. Обе точки зрения имеют подтверждения в антропологическом материале, и загадка происхождения этих рас еще ждет своего решения.
Автогенетические процессы и половой отбор
Изоляция и метисация могут приводить не только к сохранению или смешиванию антропологических признаков. Большое значение имеют также так называемые автогенетические процессы. Нет необходимости подробно разбирать их в настоящей главе, однако не упомянуть о них было бы также неверным.
Изоляция, например, может приводить к изменению генотипа популяции путем генетического дрейфа (дрейф генов). В силу разных случайностей частота генов в популяции может не направленно, но весьма значительно изменяться. Возникающие случайные мутации могут закрепляться в небольших популяциях. Так, например, независимо возникли центры светловолосости в племени нгалия в Центральной Австралии и на Соломоновых островах.
В случае заселения некой территории небольшой группой людей может проявляться так называемый эффект основателя. Какая-либо характерная особенность, имевшаяся у одного из первопоселенцев, может в последующем распространиться и стать типичной для всей группы в целом. Особенность эта может и не давать какого-либо явного преимущества для выживания группы, но распространиться в силу случайных причин, путем вышеупомянутого дрейфа генов.
Как изменение, так и сохранение расовых признаков может осуществляться путем полового отбора. Предпочтение и выбор брачного партнера осуществляется на основе двух противоречивых тенденций. С одной стороны, предпочитается брачный партнер, наиболее приближенный к усредненному варианту группы, наиболее стандартный, поскольку именно такой биологический тип прошел испытание временем и доказал свою высокую приспособленность. С другой стороны, всегда хочется лучшего, необычного и не такого, как у других. Поэтому предпочтительнее оказывается брачный партнер, в чем-то отличающийся от прочих. Тут уже начинает играть роль такое явление, как мода.
Достоверно оценить влияние, которое оказывал половой отбор на процесс расообразования пока трудно - у нас просто не хватает необходимых данных, однако сбрасывать его со счетов тоже нельзя.
Очаги расообразования:
В соответствии с гипотезой полицентризма, или полифилии, автором которой является Ф.Вайденрайх (1947), существовало четыре очага расообразования — в Европе или Передней Азии, в Африке южнее Сахары, в Восточной Азии, в Юго-Восточной Азии и на Больших Зондских островах. В Европе или Передней Азии сложился очаг расообразования, где на основе европейских и переднеазиатских неандертальцев возникли европеоиды. В Африке из африканских неандертальцев образовались негроиды, в Восточной Азии синантропы дали начало монголоидам, а в Юго-Восточной Азии и на Больших Зондских островах развитие питекантропов и яванских неандертальцев привело к формированию австралоидов. Следовательно, европеоиды, негроиды, монголоиды и австралоиды имеют свои собственные очаги расообразования. Главным в расогенезе были мутации и естественный отбор. Однако эта гипотеза вызывает возражения. Во-первых, в эволюции не известны случаи, когда бы идентичные эволюционные результаты воспроизводились несколько раз. Более того, эволюционные изменения всегда новые. Во-вторых, научных данных о том, что каждая раса обладает своим собственным очагом расообразования, не существует. В рамках гипотезы полицентризма позднее Г.Ф.Дебец (1950) и Н.Тома (I960) предложили два варианта происхождения рас. По первому варианту, очаг расообразования европеоидов и африканских негроидов существовал в Передней Азии, тогда как очаг расообразования монголоидов и австралоидов был приурочен к Восточной и Юго-Восточной Азии. Европеоиды передвигались в пределах Европейского материка и прилегающих к нему районов Передней Азии.
По второму варианту, европеоиды, африканские негроиды и австраловды составляют один ствол расообразования, тогда как азиатские монголоиды и американоиды— другой.
B соответствии с гипотезой моноцентризма, или. монофилии (Я.Я.Рогинский, 1949), которая основана на признании-общности происхождения, социально-психического развития, а так же одинакового уровня физического и умственного развития всех рас, последние возникли от одного предка, на одной территории. Но последняя измерялась многими тысячами квадратных километров Предполагается, что формирование рас произошло на территориях, Восточного Средиземноморья, Передней и, возможно Южной Азии.
16. АДАПТАЦИЯ ЧЕЛОВЕКА В РАЗЛИЧНЫХ ЭКОЛОГИЧЕСКИХ НИШАХ ЗЕМЛИ. ВЫСОКОГОРНЫЙ И АРИДНЫЙ АДАПТИВНЫЕ ТИПЫ, ХАРАКТЕРИСТИКА ПО Т.И. АЛЕКСЕЕВОЙ.
анализ закономерностей территориальной изменчивости биологических признаков в популяциях коренного населения разных географических регионов планеты выявил ряд важных особенностей, присущих современному человечеству.
Прежде всего, мы видим, что человек как биологический вид характеризуется огромной вариабельностью черт строения тела, физиологических и генетических признаков.
Эту вариабельность можно трактовать двояко:
с одной стороны, это биологическая предпосылка возможности существования человека в различных географических условиях,
с другой - отражение различных экзогенных влияний по мере расселения человечества.
Движущей силой миграционных процессов, сталкивающих человечество с новыми условиями, всегда были исторические причины, приспособление же к новым экологическим нишам осуществлялось не только с помощью культурных усовершенствований, но и в результате биологических перестроек, касающихся как функциональных, так и структурных систем организма.
Под влиянием естественной среды человеческие популяции приобретают свойства, делающие их более приспособленными к тем или иным географическим условиям жизни. Следовательно, строение тела и физиологические признаки, характерные для коренных жителей той или иной экологической ниши, можно рассматривать как реакцию, играющую приспособительную роль, в связи с воздействием комплекса окружающих условий.
Если мы обобщим картину изменчивости отдельных признаков, то увидим, что для жителей разных экологических зон Земли оказывается характерным совершенно определенное сочетанием биологических свойств:
в одних и тех же географических условиях разные по происхождению народы имеют одно и то же направление приспособительных реакций;
наоборот, в различных условиях обитания близкие в генетическом отношении группы характеризуются различными адаптивными чертами.
Такая норма биологической реакции на комплекс условий окружающей среды была названа адаптивным типом. Определенное сочетание черт строения тела и типа обмена веществ, которое входит в понятие адаптивного типа, обеспечивает состояние равновесия популяции с этой средой (Алексеева Т.И., 1977).
Региональные адаптивные типы независимы от расовой и этнической принадлежности и не представляют собой вариант крайней специализации. Они проявляются лишь в виде тенденции к изменению структурных и функциональных признаков в направлении, наиболее благоприятном для существования в определенной среде, что не препятствует возможности существования в другихэкологических нишах. И все же тенденция эта весьма ощутима - как мы отмечали, миграция того или иного адаптивного типа в новую среду проходит далеко не безболезненно (организм "настроен" на совершенно определенное сочетание экзогенных факторов).
9.3.2. Основные характеристики адаптивных типов
Итак, адаптивный тип - это норма реакции, независимо (конвергентно) возникающая в сходных условиях среды обитания, в популяциях, которые могут быть не связаны между собой генетически. Согласно этой гипотезе, достаточно четко выделяются несколько устойчивых комплексов биологических признаков.
1. Характерен повышенный уровень жирового и белкового обмена (вместе с этим - холестерина крови, гамма-глобулинов и т.п.). Все это создает высокую теплопродукцию и низкую поверхность теплоотдачи, что может рассматриваться как приспособление к ведущему фактору среды - холодовому стрессу. Для ряда признаков получены весьма высокие и достоверные коэффициенты корреляции с холодовым индексом Бодмана (до 0,6-0,8 для таких признаков, как теплопродукция и тотальные размеры тела).
Для арктических популяций характерно ускорение процессов роста, развития и старения, но и жизненный цикл человека несколько укорочен.
В целом характерно снижение вариабельности антропологических признаков по сравнению с населением умеренной зоны.
3. 4. Аридный адаптивный тип. Многие черты тропического комплекса свойственны и населению тропических пустынь: отмечается тенденция к линейности телосложения (высокий процент астеноидных форм с уплощенной грудной клеткой), развитие мускульного и жирового компонента понижено. Снижены уровни основного обмена, холестерина крови, как правило, и минерализации скелета. Наряду с этим отмечается более эффективная сосудистая регуляция потери тепла в условиях резких суточных колебаний температуры окружающей среды. Население внетропических пустынь отличается несколько большей плотностью тела (крупные размеры и вес) - это уже знакомая нам реакция на более низкие температуры среды.
5. Высокогорный адаптивный тип. В условиях высокогорья, для которого характерен недостаток кислорода (гипоксия) и понижение температуры среды, формируются такие признаки как массивность скелета и крупные размеры длинных костей (что связано с интенсивным эритропоэзом), цилиндрическая грудная клетка с высокой жизненной емкостью легких (ЖЕЛ). Характерно высокое содержание гемоглобина крови, увеличен периферический ток крови, отмечено большее число и величина капилляров.
Нетрудно убедиться в том, что эти особенности представляют собой приспособление к гипоксии, выступающей в сочетании с пониженной температурой среды.
В условиях высокогорья в целом менее интенсивно идут процессы роста и развития, позднее наступает старость, продолжительней жизненный цикл.
17. ХРОНОЛОГИЯ АДАПТИВНЫХ ТИПОВ. КОНТИНЕНТАЛЬНЫЙ И УМЕРЕННЫЙ АДАПТИВНЫЕ ТИПЫ, ХАРАКТЕРИСТИКА ПО Т.И. АЛЕКСЕЕВОЙ.
Континентальный адаптивный тип. Для жителей континентальной зоны характерны укороченные пропорции тела, уплощенная грудная клетка, в среднем повышенное жироотложение и явное увеличение массы тела (все чаще встречаются грудной и брюшной конституциональные типы). Из физиологических признаков заметно понижение содержания минеральных веществ в скелете.
Близкие черты присущи и жителям таежной зоны, но они отличаются, прежде всего, миниатюрностью и мезоморфностью сложения. Вариабельность антропологических признаков несколько выше, чем у представителей предыдущего адаптивного типа
Адаптивный тип умеренной зоны. Население умеренной зоны, безусловно, не остается нейтральным по отношению к воздей

