 2015-05-30
2015-05-30 1590
1590В последние годы вновь возродился интерес к проблеме физиологических механизмов сознания. На XXVI Международном психологическом конгрессе, состоявшемся в 1996 г. в Монреале (Канада), проблеме сознания был посвящен специальный симпозиум «Нейрофизиология сознания». Одним из ключевых направлений обсуждения явилась проблема связи сознания с высокочастотной активностью мозга на частоте гамма-колебаний (35—120 Гц). Р. Ллинас (Шпаз К.) в своем докладе «Сознание и когерентная активность мозга» определил сознание как синхронизацию нейронной активности. Он предположил, что когерентная электрическая активность достаточно большого числа нейронов создает необходимое и достаточное условие для явления сознания даже в том случае, если при этом сенсорные входы не работают, как, например, во время сновидений. Основу для такой когерентности образуют присущие нейронам электрические свойства, проявляющие себя в ритмах 40 Гц. Таким образом, идея М.Н. Ливанова, высказанная им в 50-х годах о пространственной синхронизации ритмической активности мозга как одном из механизмов обработки информации, получила новое развитие в представлении о кооперативных функциях нейронов, отражающихся в высокочастотной синфазной синхронизации нейронной активности в полосе гамма-колебаний.
Можно выделить два подхода к решению проблемы формирования субъективного образа. Одни исследователи связывают появление субъективного образа с представлением о «временном связывании» нейронов. Другая точка зрения подчеркивает роль гностических единиц в формировании субъективного образа.
Принцип «временного связывания» элементарных символов (нейронов) в более сложные структуры на основе корреляции их сигналов во времени был предложен К. Фон дер Малсбургом и В. Шнайдером (Уоп йег Ма15Ьиг§ С., 5сппеи1ег \У., 1986). В их модели организации сложной структуры (гештальта) нет места «гностическим единицам». Ансамбль возникает как констелляция корреляционно связанных элементарных символов (нейронов). Идея «временного связывания» нейронов в ансамбли через механизм общей для них высокочастотной синхронизации разделяется многими исследователями (Спс1(Р., КосЬ СЬ„ 1990; 5ш§ег \У., 1990; Спс^ Р., 1995). Р. Экхорн и др. (ЕскЬогп К., Ваиег К., Зотйап V/. е1 а!., 1988) вводят понятие пространственного «контраста синхронизации» между областями, представляющими разные объекты, что позволяет различать их — функция так называемой «пространственной сегментации».
Сторонники данной точки зрения исходят из феномена син-фазности высокочастотных потенциалов у нейронов зрительной коры кошки, избирательно возбуждающихся при восприятии одного и того же зрительного объекта. В. Зингер с коллегами из Института мозга им. Макса Планка во Франкфурте полагают, что колебания 40 Гц могут синхронизировать импульсацию нейронов, реагирующих на различные аспекты воспринимаемого эпизода, и, таким образом, представлять собой прямой нервный коррелят сознания.
Сходный эффект синхронизации биотоков на частоте гамма-колебаний обнаружен и у собак (Думенко В.Н., 1997). После выработки у животного пищевого инструментального условного рефлекса в периоды между стимулами (2—3 мин) собака демонстрировала поведение ожидания условного сигнала, принимала позу стойки и фиксировала глазами кормушку. Это состояние было очень устойчивым и сочеталось с усилением когерентных связей и уменьшением фазовых сдвигов между высокочастотными колебаниями (40-170 Гц) электрокортикограмм от разных зон коры. Автор рассматривает эту картину как отражение «внутреннего образа ситуации», который содержит знания, накопленные во время обучения. А феномен синфазности высокочастотных колебаний интерпретирует не только как коррелят «когнитивного образа», но и как сам механизм его образования («целостности»).
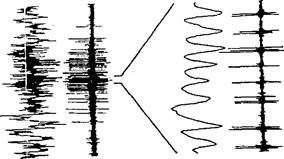
Во многих других работах также описана реакция нейронов в виде появления у них синхронизированных высокочастотных разрядов; реакция вызывается только тем стимулом, на восприятие которого нейроны избирательно настроены, например на линию, ориентированную определенным образом. Кросскорреляционный анализ ответов нейронов зрительной коры кошки, пространственно удаленных, но обладающих общей избирательностью в отношении определенных параметров стимула, показал, что такие нейроны характеризуются синхронизированными разрядами на частоте около 40 Гц. При этом фазовый сдвиг между ними равен пулю-Изменение параметров стимула может увеличить или, наоборот, уменьшить степень синхронизации этих нейронов. Так, у двух клеток зрительной коры, принадлежащих к одной группе нейронов-детекторов и находящихся на расстоянии 7 мм друг от друга, можно было видеть синхронизацию активности, когда через их рецептивные поля одинаковым образом двигались полоски (в одном направлении и с одинаковой скоростью). Если же полоски пересекали в разных направлениях рецептивные поля нейронов, синхронизация их разрядов не возникала (рис. 60). Когерентная спайковая активность зрительных нейронов обнаружена в полях 17 и 18.
В опытах на бодрствующих обезьянах была выявлена другая важная особенность феномена гамма-активности у детекторных нейронов; чем ближе стимул по своим характеристикам к селективным свойствам нейрона, тем больше у этого нейрона амплитуда локальных ритмических потенциалов на частоте 70—80 Гц. Поэтому такие ритмические осцилляции получили название стимул-специфических. С позиции модели «временного связывания нейронов» трудно объяснить этот факт. Она не может ответить также на вопрос: «Почему корреляционные функции разрядов нейронов и локальных потенциалов характеризуются нулевым сдвигом по фазе?»
Другая точка зрения на формирование субъективного образа исходит из признания иерархической организации нейронных структур. Идея принадлежит Ю. Конорскому (1970), который выдвинул концепцию гностических единиц — нейронов, реагирующих на сложные признаки в результате конвергенции на них нейронов более низкого уровня — детекторов элементарных признаков. Формирование образа связано с активацией нейронного ансамбля, который представляет собой набор нейронов, имеющих общий вход и конвергирующих на нейроне более высокого порядка. Дальнейшее развитие концепция гностических единиц получила в теории «иерархическая модель гештальта» (Соколов Е.Н., 1996). Признавая неоспоримость экспериментальных фактов — су-
Ю и •л. и Е 3
| \ |
 |
| п |
| ^ 1^ |
| ^0 |
 |
| ^ё.1 §3^ | я 5 = § ^ ^с|Й& •а е 5 ^ ЕЗ |
| р§ 5=2 % ^1 И 5 @ § Р- " 5 р " | -д* 1М та /-\ " 3 -- § § 2 ^ё в ^ И § |? § Я §1М| 0. и Х Я 3 |
| 7 " м | "^ И А Я ^ |
| я оа а- | «2 в 3 5 |
| ^ == 3 5ч-' Й 5 к "3 р| | 0 " Л Д Ч с я ° И и а = с ^11 Й1 § 1§- |
| ? и Й | 1-§1ё1 |
| а 1 ^ 2 =; з ш 1- л ~а 0 Р? Р Е СЬ & " Б. ^ с[ 1^ 1ё|^ Ё Й ^ °- •л- \и ••>* ^-< гз е1 я -ч ^ |5 И 1 0 | ч: я ь я ь «я 3 и ^ 0 и 0 &. & <у §^0§,1 1^1^ Я «1 В ё? Р^Рё 5 ё - ^ 5 3 ^|!^ §0^253 |
| Я д 5 Ю 0 Д 2 ^ § I-2 И | В м я> — -а 5 &. 0 К ^ о. ю — а ^ЕI?1 |
| 1 1^ 1 ^ §. " ё 8- ^ Е ^ 2 ^ а; и о- о " 5 \-} V (I | /~1 Т1 ^ 3 •у -б •д а " а я я 1111^ г^ь Д а Е о Я У ^ у 5 |^§1 |
| ^ а | | ^ с.1» |
| ^ с Э | и " ~ я |
| " ^3 | Т ^ К '5 ^ \ 0 v, а и |
| ^&2 | • е- о к па а и Ю о и |
ществование гностических единиц и высокочастотных когерентных колебаний между разными участками коры, Е.Н.Соколов предлагает иную интерпретации природы и механизма действия высокочастотных гамма-ритмов.
В его модели нейроны, обнаруживающие синхронизированный ритм в полосе гамма-колебаний, не связаны между собой общим источником. Но сенсорный стимул, приходящий к нейронам, одновременно возбуждает у них постсинаптические потенциалы, которые и запускают их собственную пейсмекерную активность. Если нейроны принадлежат к одной группе детекторов (например, детекторов, реагирующих на определенное направление и скорость движения), частоты их пейсмекерной активности совпадают. Такая модель объясняет увеличение амплитуды синхронизированных ритмов при приближении стимула к селективным свойствам нейрона, Пейсмекер генерирует максимальную активность, когда характеристики сенсорного сигнала совпадают с резонансной частотой данного пейсмекера. Нейроны-детекторы с пейсме-керными свойствами, принадлежащими к одной группе, имеют общую резонансную частоту. Отсутствие фазового сдвига у феномена синхронизации является следствием одновременного прихода сенсорного сигнала на нейроны-детекторы с идентичными свойствами. Таким образом, когерентность высокочастотных колебаний в группе нейронов, по-видимому, возникает в результате прихода к ним общего сигнала, на который они одинаково реагируют. Поэтому синхронизация нейронной активности на частоте гамма-колебаний не является выражением работы специального механизма связывания элементов.
Данные о пейсмекерных свойствах специфических нейронов таламуса, которые способны генерировать высокочастотные ритмы за счет высокопороговых кальциевых каналов, локализованных на их дендритах, говорят в пользу этой гипотезы. Специфический сигнал на уровне таламуса запускает особую форму несце-цифических влияний — высокочастотные ритмические колебания, частота которых, по-видимому, связана со специфичностью стимула. Такая частотно-специфическая активность выделяет лишь свой ансамбль нейронов, который избирательно настроен на восприятие данного стимула, вызывая у него особое состояние — режим синхронизированной активности.
В концепции Е.Н. Соколова механизм появления субъективного образа связан с активацией гностической единицы, которая является вершиной пирамиды, построенной из нейронных детекторов признаков. Дополнительная активация гештальт-пирамиды, поступающая от активирующей системы мозга— необходимое уело -316
| Гностический нейрон |
^з-"
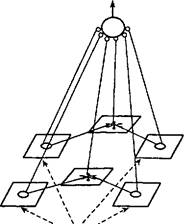
Рис. 61. Нейронная структура гештальт- пирамиды.
На нижнем уровне — простые детекторы (кружки). Следующий уровень представлен сложными детекторами (звездочки). Пунктиром показаны пути воздействия 5-стимула. Детекторы простых и сложных признаков конвергируют на гностическом нейроне (вершина пирамиды) (по Е.Н- Соколову, 1996).
вие каждого осознанного восприятия. Активирующие влияния достигают гностической единицы в форме высокочастотных колебаний мембранного потенциала. Только конвергенция специфического сигнала с механизмом неспецифической активации на нейронах таламуса создает необходимое условие для сознательного переживания. Возбуждения из таламуса сначала достигают нейронов-детекторов элементарных признаков, организованных в соответствующие карты — экраны. Элементарный признак вызывает максимум возбуждения на определенном месте этого экрана. Комплексные признаки образованы комбинациями элементарных признаков и кодируются максимумами возбуждения на картах сложных детекторов. Детекторы простых и сложных признаков образуют основание гештальт-пирамиды и имеют синапсы на гностической клетке (рис. 61). При действии сложного стимула нейроны на разных уровнях соответствующей гештальт-пирамиды «подсвечиваются» активирующими влияниями, представленными внутриклеточными колебаниями мембранного потенциала клеток. Такая активированная гештальт-пирамида иерархически организованных нейронов представляет базисный механизм актов сознания (Соколов Е.Н„ 1996).
Существуют три уровня субъективного отражения: перцептивный, мнемический и семантический. Возникший перцептивный образ в результате активации гештальт-пирам иды сличается с набором следов долговременной памяти и фиксируется в ее специальных нейронах. Нейроны памяти имеют двусторонние связи с семантическим экраном. Семантические нейроны реализуют акт категоризации. В обычных условиях все три уровня отображения слиты в едином акте сознания.








