 2015-06-16
2015-06-16 2345
2345Как известно, в современных клетках функции ДНК заключаются в получении, хранении и передаче информации последующим поколениям. Без ДНК и РНК невозможно точное воспроизведение всех свойств клетки, в основе которых лежит функционирование специфических белков. В модельных опытах была показана относительная простота и легкость возникновения пространственно обособленных систем, построенных из протеиноидов, характеризовавшихся определенным постоянством аминокислотных последовательностей. Это могло служить указанием на то, что информация о полипептидах типа протеиноидов была заключена в них самих, а следовательно, подводило к следующему выводу: на начальном этапе эволюции протоклетки могли воспроизводиться и передавать информацию потомству без участия нуклеиновых кислот.
Дальнейшее усложнение структуры и совершенствование функции полипептидов приводило к появлению в них определенных аминокислотных группировок, которым в какой-то степени была присуща полезная для протоклетки каталитическая активность. Однако возникновение более "совершенного" полипептида создавало преимущество для породившей его протоклетки только в том случае, если появившееся определенное сочетание аминокислотных остатков в полипептиде могло быть передано дочерним протоклеткам. При отсутствии такой способности возникшее "удачное" сочетание аминокислотных остатков в полипептиде терялось при последующем разрастании протоклеток. Таким образом, для дальнейшей эволюции протоклеток необходимо было создание специального аппарата, который обеспечивал бы в ряду их поколений точное воспроизведение полипептидов с определенно закрепленным расположением аминокислотных остатков. Это привело к формированию принципиально нового механизма синтеза — матричного синтеза, в основе которого лежит использование свойств нового класса органических соединений — полинуклеотидов.
Свойством полинуклеотидов, сформированных из одного типа асимметрических молекул, является способность к точному воспроизведению, основанная на принципе структурной комплементарности. В модельных опытах было показано, что поли- нуклеотидная цепь может служить матрицей, связывающей свободные нуклеотиды. При смешивании АМФ с полиуридиловой кислотой свободные молекулы АМФ связываются с остатками полиуридиловой кислоты при помощи водородных связей между комплементарными основаниями. В результате возникала спиральная структура. Точно так же наблюдали формирование устойчивой комплементарной спирали при смешивании полицитидиловой кислоты с гуанозинмонофосфатом. Для синтеза комплементарных полинуклеотидов необходимо было, чтобы между связанными с матрицей мононуклеотидами образовались межнуклеотидные связи. Экспериментально была показана принципиальная возможность возникновения таких связей без какого-либо участия ферментов. Таким образом, полинуклеотиды могли служить матрицей для неферментативного синтеза комплементарных полинуклеотидов.
Вопрос о том, каким путем в молекулах полинуклеотидов возникла и закрепилась информация о структуре белков, остается наиболее неясным. Имеются данные об избирательном взаимодействии между двумя типами полимеров — полиаминокислотами и полинуклеотидами — в зависимости от их аминокислотного и нуклеотидного составов, на основании чего высказывается предположение, что в принципе полиаминокислоты и полинуклеотиды могли "узнавать" друг друга в протоклетках. Образование специфических комплексов между этими полимерами можно рассматривать как первый необходимый шаг на пути установления между ними определенных "информативных" связей. Не исключено также, что на первых этапах поток информации шел в любом направлении (полинуклеотид протобелок) и, таким образом, устанавливались взаимные связи между определенными последовательностями аминокислот в протобелках и нуклеотидов в полинуклеотидах. Позднее поток информации стал однонаправленным (полинуклеотид протобелок).
Таким образом, дискуссионным остается вопрос о том, на каком этапе эволюционного процесса нуклеиновые кислоты сформировались как информационные молекулы. Согласно одним представлениям на начальном этапе эволюции роль последних выполняли белковоподобные молекулы, и первые примитивные клетки функционировали без нуклеиновых кислот. Другая гипотеза исходит из того, что первыми возникли нуклеиновые кислоты, а позднее, на базе содержащейся в них информации, возникли белки (гипотеза "генной жизни").
Эта гипотеза принадлежит американскому генетику Г. Мёллеру (Н. Muller), высказавшему предположение, что "жизнь" началась с абио-генного образования гена или группы генов. Появление мембран и белков, обладающих каталитическими свойствами, имело место на более поздних этапах. В пользу этой гипотезы приводятся соображения, первое из которых основано на современном представлении о молекулярной структуре и самовоспроизведении вирусов, а второе — на полифункциональных свойствах мононуклеотидов. Хорошо известно, что нуклеотиды, помимо того, что составляют генетический аппарат клетки, принимают участие в разнообразных метаболических реакциях: служат переносчиками энергии (АДФ, АТФ), электронов и атомов водорода (НАД, НАДФ, ФМН, ФАД), сахаров, ацильных групп и др.
Формы жизни, возникшие на "белковой основе", были неустойчивыми из-за отсутствия системы передачи информации, использующей свойства нуклеиновых кислот, а "генная жизнь" не могла прогрессивно эволюционировать без участия белков, обладающих каталитическими свойствами. Как произошло возникновение формы жизни, в основе которой лежат белки и нуклеиновые кислоты, пока не известно. Ясно только, что "встреча" обоих типов соединений положила начало пути эволюции, на котором произошло формирование механизмов синтеза белка и нуклеиновых кислот и кодовых взаимодействий между обоими механизмами.
ДАННЫЕ ПАЛЕОНТОЛОГИИ О ПРОИСХОЖДЕНИИ ЖИЗНИ НА ЗЕМЛЕ
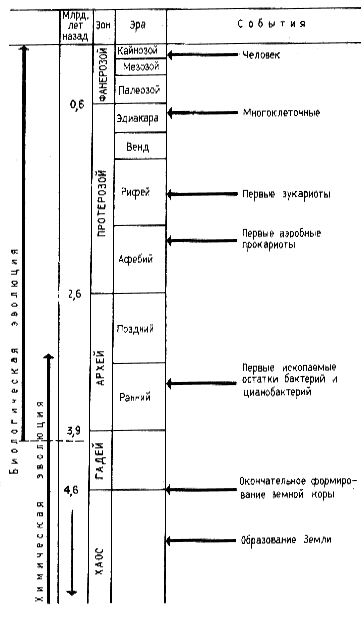 |
| Рис. 52. Этапы биологической эволюции (по Опарину, 1976; Fox, Dosel, 1975; Lehninger, 1974) |
Согласно современным представлениям окончательное формирование земной коры произошло около 4,6 млрд лет назад. Наши сведения об истории возникновения и развития жизни на Земле ограничены преимущественно последним периодом, длительность которого порядка 600 млн лет39. Остальной временной период, составляющий примерно 90% всей истории существования Земли, фактически является чистой страницей в изучении возникновения и развития жизни на Земле. Поэтому большой интерес представляют данные молекулярной палеонтологии изучающей органические вещества древнейших осадочных отложений. Трудность заключается в интерпретации полученных результатов, т. е. в отсутствии надежных критериев на основании которых можно было бы делать выводы о происхождении обнаруженных органических остатков: биогенном или абиогенном. В этой связи интересны находки, сделанные в Южной Африке в осадочных породах, возраст которых составляет около 3,5 млрд лет. В этих породах найдены заключенные в них окаменелые остатки палочковидных структур размером 05x025 мкм, напоминающих современные бактерии. При электронно-микроскопическом изучении у них выявлена двухслойная клеточная стенка, подобная клеточной стенке многих современных бактерий.
39 В геологии (см. рис. 52) приняты следующие названия временных интервалов: зон, эра, период, эпоха, век. Самое крупное геохронологическое образование — зон. Зоны подразделяются на эры и т. д. Фанерозой, самый последний и наиболее изученный зон, включает три эры: палеозой, мезозой и кайнозой Первый период палеозойской эры — кембрий (начало примерно 580 млн лет назад, длительность — 80 млн лет). Характеризуется появлением животных, имеющих скелетные элементы. Более ранние зоны часто объединяются под названием докембрия или дофанерозоя.
В породах, возраст которых также около 3, 5 млрд лет, обнаружены строматолиты, своеобразные известковые образования, являющиеся продуктами жизнедеятельности древних фотосинтезирующих организмов — цианобактерий, или синезеленых водорослей. Если принять, что найденные в породах ископаемые остатки действительно принадлежат древнейшим прокариотам или являются продуктами их жизнедеятельности, то следует признать, что к этому времени уже были сформированы некоторые типы жизни, которые дошли до нас в виде ее "следов" Отсюда приходится сделать вывод, что впервые земная жизнь должна была возникнуть в промежутке между 3,5 и 4,6 млрд лет тому назад, однако у нас нет никакой информации об этом периоде. Схематическое изображение во времени отдельных этапов эволюции представлено на рис. 52.
Цианобактериям мы обязаны появлением молекулярного кислорода в атмосфере Земли. Однако вначале весь выделяемый ими Оа поглощался земной корой, в которой происходили интенсивные процессы окисления. По имеющимся геологическим данным содержание кислорода в атмосфере достигло 1% от его содержания в современной атмосфере только в среднем протерозое, и к этому времени можно отнести возникновение первых аэробных прокариот. В пользу этого свидетельствуют обнаруженные в отложениях, возраст которых около 2 млрд лет, звездчатые образования, свойственные облигатно аэробной свободноживущей бактерии Metallogenium. Этот организм откладывает на поверхности клеток окислы железа. В природе встречается при разных концентрациях O2, но всегда в аэробных условиях, так что может служить индикатором молекулярного кислорода.
Первые эукариоты появились приблизительно 1,5 млрд лет назад. Таким образом, прокариоты были единственными обитателями нашей планеты в течение 2/3 времени эволюции биосферы. Жизнедеятельность прокариот привела к накоплению в атмосфере молекулярного кислорода и к обогащению лито- и гидросферы органическим веществом.
 |  |  |  |  |
| Назад | Главная страница | Оглавление | Вниз | Вперед |
III. Эволюция энергетических процессов у эубактерий
В главах 11 и 12 были обсуждены проблемы возникновения первичной клетки из гипотетической протоклетки и последующего пути прогрессивной эволюции первичной клетки. Как было обнаружено в 70-х гг., на раннем этапе этого пути могло произойти выделение трех основных ветвей, каждая из которых самостоятельно и независимо эволюционировала, результатом чего явилось существование в рамках прокариотной клеточной организации двух крупных таксономических групп: эубактерий и архебактерий.
Подавляющее большинство известных прокариот относится к эубактериям. Именно у них достаточно хорошо изучены основные этапы прогрессивной эволюции, связанные с совершенствованием энергетических процессов. Архебактерии представляют собой к настоящему времени разрозненные группы со специфическими типами энергетического метаболизма. Имеющегося материала явно не достаточно для создания каких-либо эволюционных построений внутри группы архебактерий.
ГЛАВА 13. БРОЖЕНИЕ. ТИПЫ ЖИЗНИ, ОСНОВАННЫЕ НА СУБСТРАТНОМ ФОСФОРИЛИРОВАНИИ
Наиболее примитивным способом получения энергии, присущим определенным группам эубактерий, являются процессы брожения.
ОБЩАЯ ХАРАКТЕРИСТИКА ПРОЦЕСCOВ БРОЖЕНИЯ
"Брожение" — это сугубо микробиологический термин. Он характеризует энергетическую сторону способа существования нескольких групп эубактерий, при котором они осуществляют в анаэробных условиях окислительно-восстановительные превращения органических соединений, сопровождающиеся выходом энергии, которую эти организмы используют. Поскольку брожение протекает без участия молекулярного кислорода, все окислительно-восстановительные превращения субстрата происходят за счет его "внутренних" возможностей. Процесс брожения связан с такими перестройками органических молекул субстрата, в результате которых на окислительных этапах процесса высвобождается часть свободной энергии, заключенной в молекуле субстрата, и происходит ее запасание в молекулах АТФ. В процессе брожения, как правило, происходит расщепление углеродного скелета молекулы субстрата.
Круг органических соединений, которые могут сбраживаться, довольно широк. Это углеводы, спирты, органические кислоты, аминокислоты, пурины, пиримидины. Химическое вещество может быть подвергнуто сбраживанию, если оно содержит неполностью окисленные (или восстановленные) углеродные атомы. В этом случае есть возможность для окислительно-восстановительных преобразований между молекулами (или внутри одного вида молекул), возникающими из субстрата. В результате одна часть продуктов брожения будет более восстановленной, другая — более окисленной по сравнению с субстратом. Продуктами брожений являются различные органические кислоты (молочная, масляная, уксусная, муравьиная), спирты (этиловый, бутиловый, пропиловый), ацетон, а также CO2 и H2. Обычно в процессе брожения образуется несколько продуктов. В зависимости от того, какой основной продукт накапливается в среде, различают молочнокислое, спиртовое, маслянокислое, пропионовокислое и другие виды брожений.
Следовательно, в каждом виде брожения можно выделить две стороны: окислительную и восстановительную. Процессы окисления сводятся к отрыву электронов от определенных метаболитов с помощью специфических ферментов (дегидрогеназ) и акцептированию их другими молекулами, образующимися из сбраживаемого субстрата, т. е. в процессе брожения происходит окисление анаэробного типа.

