 2015-06-28
2015-06-28 3510
3510Матричные синтезы
Удвоение молекулы ДНК – репликация. В результате этого и последующего деления дочерние клетки наследуют геном родителей, в котором полный набор генов, или инструкция о строении РНК и всех белков организма. Это первый поток передачи информации.
Второй поток – происходит в процессе жизнедеятельности клетки. Происходит считывание, или транскрипция, генов в форме полинуклеотидных последовательностей мРНК и использование их как матрица для синтеза соответствующих белков. Т.е. происходит перевод, или трансляция информации с мРНК на язык аминокислот. Поток информации от ДНК через РНК на белок – центральная догма биологии.
Исправление ошибок в структуре ДНК, возникающих под действием внешних и внутренних факторов, осуществляет еще один матричный синтез – репарация.
Итак, к матричным синтезам относят репликацию, транскрипцию, трансляцию и репарацию.
Репликация
Хромосома содержит одну непрерывную двухцепочечную молекулу ДНК. При репликации каждая цепь родительской ДНК служит матрицей для синтеза новой комплементарной цепи. Вновь образованная двойная спираль имеет одну исходную и одну вновь синтезируемую цепь. Такой механизм носит название полуконсервативная репликация.
Репликация состоит из стадий:
Инициация – образование репликативной вилки
Элонгация – синтез новых цепей
Исключение праймеров
Терминация
Синтез ДНК происходит в S-фазу. Инициацию репликации регулируют специфические сигнальные белки – факторы роста. Они связываются с рецепторами мембран, передающих сигнал, побуждающий клетку к началу репликации. Синтез новых одноцепочечных молекул ДНК может произойти только при расхождении родительских цепей. В определенном сайте (точка начала репликации) происходит локальная денатурация ДНК, цепи расходятся и образуются две репликативные вилки, движущиеся в противоположных направлениях. Образование репликативной вилки:
ДНК-топоизомеразы (I, II, III) обладают нуклеазной активностью. ДНК-топоизомераза I разрывает фосфоэфирную связь в одной из из цепей и ковалентно присоединяется к 5’-концу в точке разрыва.По окончании формирования репликативной вилки фермент ликвидирует разрыв и отделяется от ДНК. Разрыв водородных связей в двухцепочечной ДНК осуществляет ДНК-хеликаза. Она использует АТФ для расплетения двойной спирали. В результате происходит раскручивание участков суперспирализованной молекулы. В поддержании этого участка в раскрученном состоянии участвуют SSВ-белки. Кготорые связываются с одноцепочечной нитью. Эти белки не закрывают азотистые основания, но не дают комплементарное скручивание и образование шпилек. Они обладают большим сродством к одноцепочечным участкам.
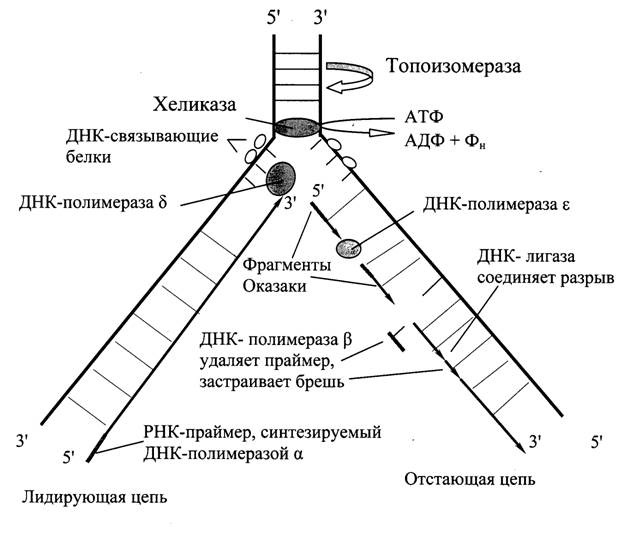
Репликация осуществляется ДНК-полимеразами. Субстратами и источниками энергии служат дезоксирибонуклеозидфосфаты дАТФ, дГТФ, дЦТФ и дТТФ. Для их активации необходимы ионы магния, т.к. они нейтрализуют отрицательный заряд и повышают их реакционную способность. Синтез происходит в направлении 5’ → 3’ растущей цепи, т.е. очередной нуклеотид присоединяется к свободному 3’-ОН- концу предшествующего нуклеотида. Синтезируемая цепь всегда антипараллельна матричной цепи. В ходе репликации образуются 2 дочерние цепи, являющиеся копиями матричных цепей.
Существует 5 ДНК-полимераз (α, β, γ, δ, ε). Они различаются по числу суъединиц, молекулярной массе и функциональному назначению. ДНК-полимеразы α, β, δ, ε участвуют в синтезе ДНК в ядре, а γ в репликации митохондриальной ДНК.
Репликацию инициирует ДНК-полимераза α, т.к. коплементарна определенному сайту одноцепочечной ДНК. Она синтезирует небольшой фрагмент РНК – праймер, состоящий из 8-10 рибонуклеотидов. Далее она синтезирует олигонуклеотид их ≈ 60 нуклеотидных остатков. Первые 8-10 представлены рибонуклеотидами, остальные – дезоксирибонуклеотидами.
Этот олигонуклеотид, образующий небольшой двухцепочечный фрагмент с матрицей позволяет присоединиться ДНК-полимеразе δ и продолжить синтез новой цепи в направлении от 5’- к 3’- концу по ходу раскручивания репликативной вилки. В каждой репликативной вилке идет одновременно синтез двух новых цепей. Лишь для одной цепи совпадает движение с репликативной вилкой – это лидирующая цепь. Для другой цепи синтез осуществляется ДНК-полимеразой α и ε в направлении 5’- к 3’- концу, но против движения репликативной вилки. Поэтому вторая цепь синтезируется прерывисто, короткими фрагментами (фрагменты Оказаки). Эту цепь называют отстающей. Каждый фрагмент ≈ 100 нуклеотидови содержит праймер. Праймеры удаляет ДНК-полимераза β, постепенно отщепляя по одному рибонуклеотиду и присоединяет к ОН-группе на 3’- конце дезоксирибонуклеотиды. Далее ДНК-лигаза закрывает брешь и образуется непрерывная цепь ДНК.
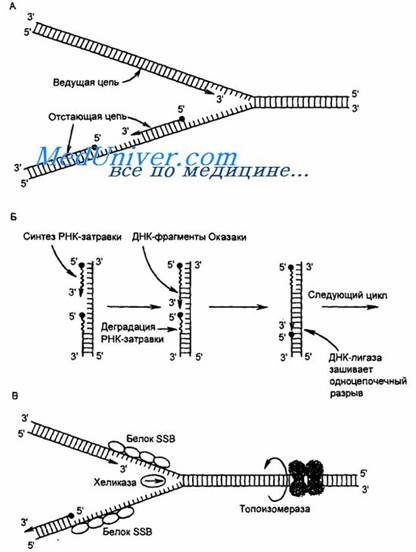
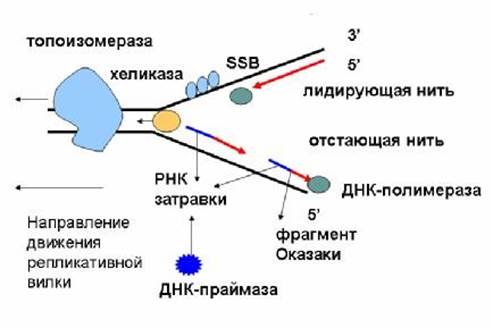
Инициация ДНК происходит в нескольких сайтах хромосомы. Их называют сайтами репликации, или ориджинами. Последовательность ДНК, ограниченную двумя ориджинами, называют единицей репликации, или репликоном. Две репликативные вилки двигаются в противоположных направлениях до тех пор пока не встретяться.
После завершения репликации происходит метилирование нуклеотидных остатков вновь образованных цепей. Метильные группы присоединяются ко всем остаткам аденина в последовательности GATC с образованием
N6-метиладенин или возможно метилирование цитозина в последовательности GC с образованием N5-метилцитозина. Количество метилированных оснований равно ≈ 1-8%. Модификация происходит при участии ферментов. Использующих метильные группы S-аденозилметионина (SАМ). Присоединение метильных групп к остаткам аденина и цитозина не нарушает комплементарности цепей.
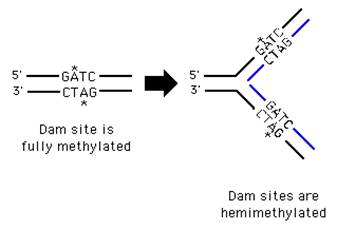
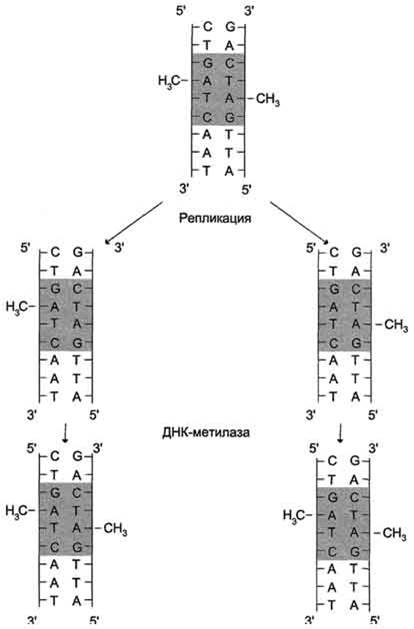
Наличие метильных групп в цепях ДНК необходимо для формирования структуры хромосом и для регуляции транскрипции генов.
В течение непродолжительного времени в молекуле ДНК последовательности – GATC- метилированы по аденину только в матричной цепи. Это различие используется ферментами репарации для исправления ошибок репликации.
На каждом конце хромосомы присутствует специфическая нуклеотидная последовательность. Она представлена многочисленными повторами (сотни или тысячи раз) олигонуклеотидов – GGGTTA-. Это сочетание называют теломерной последовательностью, или теломерной ДНК. Наличие теломер необходимо для завершения репликации концевых информативных последовательностей хромосом, т.е. для сохранения генетической информации. После завершения репликации хромосомы 5’-конца дочерних цепей ДНК недостроены, т.к. после удаления праймеров эти фрагменты оказываются недореплицированными, потому что ДНК-полимераза β, ответственная за заполнение бреши не может вести синтез цепи ДНК от
3’- к 5’- концу. Таким образом в ходе каждого цикла репликации 5’- концы синтезированных цепей укорачиваются. Эти потери не представляют опасности для генетической информации, т.к. укорочение идет за счет теломер.
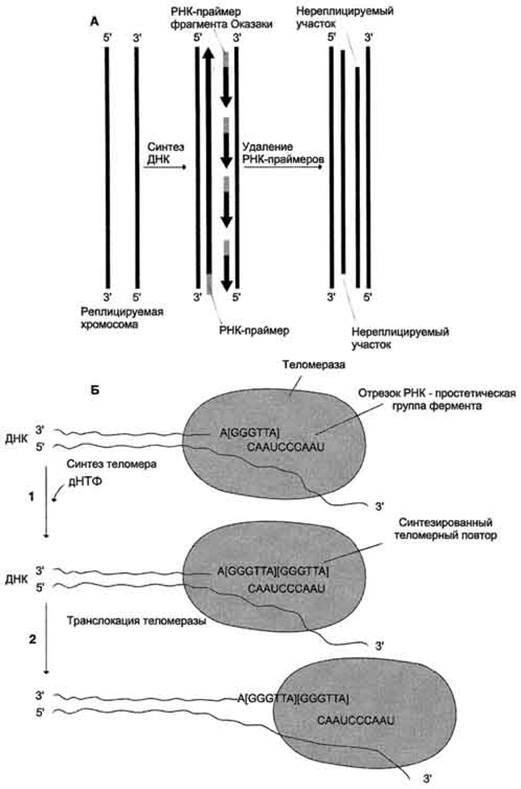
Т.о. с каждым клеточным делением ДНК хромосом будут последовательно укорачиваться. Укорочении теломер в большинстве клеток по мере их старения – важный фактор, определяющий продолжительность жизни организма.
Однако в эмбриональных и других быстро делящихся клетках потери концов хромосом недопустимы. В клетках имеется фермент теломераза (нуклеотидилтрансфераза), которая восстанавливает недореплецированные
5’-концы. В ферменте в качестве простетической группы присутствует РНК. Она находится в активном центре фермента и служит матрицей при синтезе теломерных повторов хромосом, т.е. постепенно наращивает гексануклеотид – GGGTTA-. В большинстве клеток она не активна. Однако небольшая ее активность обнаруживается в лимфоцитах, стволовых клетках костного мозга. клетках эпителия, эпидермисе кожи.

