 2015-06-28
2015-06-28 2575
2575Мышечная двигательная активность практически непрерывно сопровождает все проявления жизнедеятельности человека. Это совершенно понятно, когда речь идет о каких-либо физических упражнениях, как бытовых, так и специальных. Но не только в таких условиях. Когда человек спокойно стоит, сидит и даже лежит, его скелетная мускулатура не приходит в состояние полного покоя. Ведь каждое из названных положений представляет определенную позу, которая направлена на противодействие силе земного притяжения. Более того, даже в состоянии глубокого естественного сна не происходит полного расслабления мышечного аппарата человека.
Сопровождается ли мышечная активность какими-либо специфическими ощущениями? Не торопитесь с ответом. Как это принято в физиологии, попытаемся ответить на этот вопрос экспериментально. Попросите вашего соседа закрыть глаза. А затем придайте его руке любое положение. Для наглядности лучше такое, чтобы участвовали все суставы. Затем попросите этого человека, чтобы он, не открывая глаз, теперь уже самостоятельно придал второй руке такое же положение. И вы убедитесь, что это задание будет выполнено быстро, с большой точностью и без каких-либо затруднений. Этот немудреный опыт порождает очень сложный вопрос: «Как правая рука ведает, что делает левая?»
Давайте теперь проанализируем факт, который хорошо знаком каждому из повседневной жизни. Наверное, не раз случалось, находясь в неудобной позе, «отсидеть» или «отлежать» ногу либо руку. Такое состояние всегда сопровождается временным, полным или частичным нарушением чувствительности.
Обратите внимание — нарушением чувствительности. Вспомните, сколь не точными при этом становятся движения такой конечности, и совершенно не удается продублировать ее положение на противоположной стороне без контроля глаз. А если вы никогда не обращали на такое явление внимания, то при первом удобном случае попытайтесь это проверить. Из рассмотренных, в общем-то, широко известных фактов логичным будет сделать, по крайней мере, два предположения. Во-первых, наши мышцы, или, точнее, опорно-двигательный аппарат, наделены чувствительностью. И, во-вторых, такой вид чувствительности необходим для координации мышечной деятельности.
Эти предположения, к которым мы пришли, анализируя наши повседневные наблюдения, явились предметом весьма многочисленных исследований. К настоящему времени накоплено много и морфологических, и функциональных данных, позволяющих говорить о двигательном анализаторе как о совокупности нервно-рецепторных образований, которые воспринимают состояние опорно-двигательного аппарата и обеспечивают формирование соответствующих ощущений, сопровождающихся двигательными и вегетативными рефлексами. Иными словами говоря, биологическая роль двигательного анализатора заключается в обеспечении координации двигательной активности и снабжении работающих мышц необходимыми веществами.
Нервные окончания в структурах опорно-двигательного аппарата весьма разнообразны по форме и механизмам функционирования. Они располагаются в мышцах, сухожилиях, фасциях, надкостнице, тканях суставов. Здесь можно обнаружить рецепторные образования, встречающиеся и в других частях тела (в частности, такие, какие рассматривались при описании тактильной и температурной чувствительности), а также и специализированные чувствительные структуры, присущие только двигательному анализатору. Их часто называют проприоцепторам или проприорецепторами, а обусловленную ими чувствительность как проприоцептивную (проприорецептивную) чувствительность. Такими специфическими рецепторами опорно-двигательного аппарата являются сухожильные органы Гольджи и мышечные веретена. Оба вида чувствительных образований по механизму функционирования относятся к механорецепторам, т. е. воспринимающим механическую энергию, однако их конкретная роль в передаче информации неоднозначна.
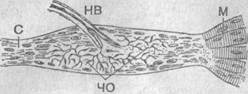
Рис. 29. Сухожильный орган Гольджи:
С — сухожилие; НВ — чувствительное нервное волокно; Ч0 — его чувствительные окончания; М — мышечные волокна.
Сухожильные органы Гольджи (описаны в 1880 г. выдающимся итальянским гистологом, лауреатом Нобелевской премии К. Гольджи) располагаются в сухожилиях обычно на границе мышечной и сухожильной ткани, в опорных участках капсул суставов, в суставных связках (рис. 29).
Из рисунка понятно, что это рецепторное образование расположено «последовательно» (по аналогии с электротехническими схемами) в цепи «мышца — сухожилие». Отсюда следует, что раздражение данного рецептора развивается тогда, когда возникает растяжение в этой цепи. Это, в частности, отмечается при наличии даже небольшого сокращения мышцы, т. е. даже в состоянии покоя. И степень возбуждения рецептора будет тем сильнее и тем значительнее, чем интенсивнее сокращение. Кроме того, при приложении какой-то внешней силы, растягивающей эту систему (масса самой мышцы, конечности), возбуждение в рецепторах также возрастает.
В естественных условиях, таким образом, аппарат Гольджи никогда не бывает в покое, но степень его возбуждения отражает интенсивность растяжения структуры, в которой он расположен. Для многих ситуаций такая способность является вполне достаточной, чтобы посылать в центральную нервную систему информацию, отражающую состояние опорно-двигательного аппарата.
Представим себе несколько абстрактную, но по своей принципиальной сущности реальную ситуацию (рис. 30). На этой схеме показаны контуры костей коленного сустава человека, вычерченные на основании рентгенограмм. Пунктирные линии а, б и в соединяют произвольно выбранные точки, между которыми могут быть натянуты связки.
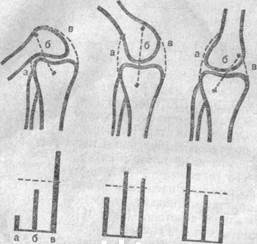
Рис. 30. Функционирование рецепторов при различных положениях сустава.
Длина черных столбиков пропорциональна длине связок а, б и в. Если представить, что в этих связках расположены рецепторы растяжения типа органа Гольджи, то тогда можно считать, что длина столбиков передает степень возбуждения рецепторов, заключенных в этих связках, а пунктирная горизонтальная линия — порог появления афферентной импульсации. Из этого следует, что в первом примере (максимальное сгибание) надпороговое возбуждение возникает только в рецепторах связки в, во втором примере (промежуточное положение) — в б и в, а в третьем (максимальное разгибание) — только в рецепторах связки а. Таким образом, в каждом разобранном случае и во всех промежуточных не рассмотренных, для всякого определенного положения суставов существует своя специфическая картина афферентной импульсации, поступающей от рецепторных структур в центры двигательного анализатора.
Вторым видом специфических рецепторных образований опорно-двигательного аппарата являются так называемые мышечные веретена, описанные еще в середине прошлого столетия. Они представляют собой вытянутые структуры, расширенные в середине за счет капсулы и напоминающие по форме веретена.
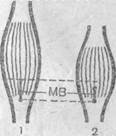 Рис. 31. Изменение степени растяжения мышечного веретена (МВ) при расслаблении (1) и сокращении (2) мышцы.
Рис. 31. Изменение степени растяжения мышечного веретена (МВ) при расслаблении (1) и сокращении (2) мышцы.
В отличие от органа Гольджи, расположенного «последовательно» между мышцей и сухожилием, мышечное веретено в этой цепи расположено «параллельно». Это и определяет специфику условий, при которых возбуждается такой рецептор. Непосредственной причиной возбуждения мышечного веретена и в этом случае является его растяжение. А теперь давайте постараемся представить, при каком состоянии мышцы будет растянуто мышечное веретено (рис. 31). Легко понять, что при сокращении мышцы точки прикрепления мышечного веретена сближаются, а при расслаблении — удаляются, т. е. мышечное веретено растягивается. Из этого следует, что данные рецепторные структуры возбуждаются во время расслабления мышцы, а степень их возбуждения будет пропорциональна степени расслабления. Мышечное веретено по своим физическим свойствам весьма эластичное образование, вследствие чего даже при реально возможных максимальных сокращениях сохраняется некоторая степень его растяжения и, следовательно, некоторая степень его возбуждения. Легко догадаться, что при искусственном механическом растяжении сухожильно-мышечной структуры в мышечном веретене, так же как и в органе Гольжди, возбуждение будет усиливаться.
Наличие таких двух рецепторных образований позволяет получать тонко дифференцированную информацию о состоянии мышцы, т. е. степени ее сокращения, расслабления или растяжения. При расслабленном состоянии мышцы имеется редкая тоническая афферентная импульсация от сухожильных рецепторов Гольджи и усиленная от мышечных веретен. При сокращении отмечается противоположное соотношение. При искусственном растяжении афферентация усиливается от обоих видов рецепторов. Таким образом, любое состояние мышцы находит отражение в характере импульсации от обоих видов рецепторов сухожильно-мышечных структур.
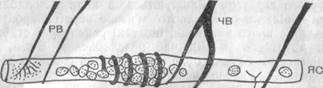
Рис. 32. Схема строения и иннервации двух типов (ЯС и ЯЦ) интрафузальных мышечных волокон:
ЧВ — чувствительные нервные волокна; ДВ — двигательные нервные волокна; 1,,2 — первичные и вторичные нервные окончания.
Рассмотрим более детально структуру и свойства мышечного веретена. Каждое мышечное веретено состоит, как правило, из нескольких так называемых интрафузальных мышечных волокон, в которых различают центральную часть и периферическую — мионевральную — трубку. Существует два типа интрафузальных мышечных волокон: ЯС-волокна, у которых ядра сконцентрированы в центральной части в виде ядерной сумки, и ЯЦ-волокна с расположением ядер в виде ядерной цепочки (рис. 32).
Количество мышечных веретен и содержание в них интрафузальных мышечных волокон в различных мышцах неодинаково. Можно заметить, что, чем сложнее и тоньше выполняемая мышцей работа, тем больше в ней рецепторных образований. Полагают, что ЯЦ-волокна связаны с тонко координированной мышечной работой.
Интрафузальные мышечные волокна получают как чувствительную, так и двигательную иннервацию. Окончания чувствительных нервных волокон или оплетают в виде спирали центральную часть (первичные окончания), или располагаются в области миотрубки (вторичные окончания). Именно в этих нервных структурах и возникает афферентная импульсация, передаваемая в центральную нервную систему, в зависимости от степени растяжения волокна.
А какова же функция двигательных волокон, подходящих к этим рецепторным структурам? Их роль была раскрыта сравнительно недавно известным современным физиологом, шведским ученым, Нобелевским лауреатом Р.Гранитом. Дело заключается в том, что периферическая, мионевральная часть интрафузального мышечного волокна содержит сократительные элементы, состоящие из поперечнополосатых мышечных волокон (т. е. таких же, как и в обычных скелетных мышцах). При их сокращении длина интрафузального мышечного волокна, естественно, уменьшается. А если мы теперь снова обратимся к рисунку 31, то станет очевидным, что такое состояние мышечного веретена сделает его более чувствительным к расслаблению мышцы; таким образом, при помощи этих двигательных нервных волокон регулируется чувствительность мышечных веретен.
Всем хорошо понятно, сколь велик мышечный аппарат человека. Соответственно этому столь же широко распространены рецепторные структуры. Нередко, подходящие к ним чувствительные нервные волокна идут вместе с двигательными в составе нервов, которые иногда не совсем верно обозначают как двигательные. Практически все нервы смешанные, т. е. содержат как двигательные, тек и чувствительные волокна.
Чисто сенсорный путь имеет переключение в продолговатом мозге, в зрительном бугре и заканчивается в коре головного мозга. Интересно заметить, что у человека корковое представительство двигательного анализатора (т.е. чувствительная система) совпадает с корковыми двигательными структурами — передней центральной извилиной. Однако чувствительные пути идут также в соматосенсорную зону (задняя центральная извилина) и префронтальную кору. Все эти участки имеют непосредственное отношение к регуляции двигательной активности.
Помимо рассмотренного специфического сенсорного пути, проприоцептивная импульсация попадает также в мозжечок, ретикулярную формацию, гипоталамус и некоторые.другие структуры. Эти связи являются отражением роли данной импульсации в регуляции двигательной активности и деятельности внутренних органов. Последнее положение не должно вызывать удивления. Ведь всякая двигательная активность требует резкой интенсификации доставки кислорода, питательных веществ, удаления углекислого газа и других продуктов обмена веществ. А для этого необходимо усиление деятельности практически всех систем внутренних органов — кровообращения, дыхания, выделения и др. Такая согласованность станет возможной, если в вегетативные центры (которые регулируют работу внутренних органов) будет поступать информация о состоянии мышц.
Рассмотрим чисто сенсорную характеристику деятельности двигательного анализатора. Абсолютную чувствительность этой афферентной системы измерить достаточно трудно. Принято судить о ней по некоторым косвенным признакам, в частности по точности воспроизведения положения сустава и ощущению изменения его положения. Установлено, в частности, что самым чувствительным в таком смысле является плечевой сустав. Для него порог восприятия смещения при скорости 0,3° в секунду составляет 0,22—0,42°. Наименее чувствительным оказался голеностопный сустав, у него порог составляет 1,15—1,30°. Для многих суставов человек с закрытыми глазами через 10—15 секунд воспроизводит положение с ошибкой около 3 %.
Иногда для оценки чувствительности, в частности, дифференциальной, двигательного анализатора используют величину едва различимой разницы в силе тяжести. В весьма широком диапазоне исследуемых значений эта величина близка к 3 %.
Адаптация в двигательном анализаторе на рецепторном уровне выражена слабо. Благодаря этому афферентная импульсация длительное время не изменяется при неизменной степени растяжения рецепторов. Однако интегральная чувствительность сенсорной системы в целом меняется в зависимости от нагрузки на опорно-двигательный аппарат. Хорошо известна его тренируемость, что выражается в развитии очень тонкой двигательной координации соответствующих групп мышц у ювелиров, музыкантов, хирургов и т. п.
С полным основанием можно говорить об исключительной значимости двигательного анализатора в выработке у человека пространственных представлений о внешнем мире. Проприоцепция для человека является основой, можно даже сказать, абсолютным критерием удаленности и размеров предмета. Ведь действительно, чтобы сложилось первоначальное представление о расстоянии до предмета, его габаритах, необходимо это расстояние «отмерить» во время ходьбы или дотянуться до предмета рукой и ощупать его. Неоднократные сочетания такого рода ощущений со зрительными, слуховыми, тактильными ощущениями позволяют выработать способность оценивать расстояния и размеры только на основе работы зрительного, слухового, кожного анализаторов. Механизмы таких ощущений, естественно, имеют свои особенности, которые и рассматривались в соответствующих главах.
Постоянной и плохо восполнимой функцией двигательного анализатора является его участие в рефлекторном формировании мышечного тонуса. Человек всегда (за исключением условий космического полета) находится под воздействием силы земного притяжения. Под ее влиянием голова, туловище, конечности и суставы принимают определенное положение, а мышцы подвергаются некоторой степени растяжения. Все это, естественно, сопровождается раздражением рецепторов мышц, сухожилий, суставных структур. Отсюда следует, что от них в центральную нервную систему постоянно поступает афферентная импульсация той или иной интенсивности, а в ответ на нее рефлекторно поддерживается соответствующая степень тонического сокращения всех скелетных мышц. Такой тонус, с одной стороны, является базисом, на котором развиваются сокращения, а с другой, обеспечивает поддержание той или иной адекватной позы.
Жизнь человека невозможно представить без движений. Двигательный анализатор — одно из звеньев управления двигательной активностью. Очень точно биологическое значение двигательного анализатора оценил И. М. Сеченов (1891): «Мышечное чувство может называться ближайшим регулятором движений и в то же время чувством, которое помогает животному познавать в каждый данный момент положение в пространстве, при том как при покое его, так и при движении. Оно представляет, следовательно, одно из орудий ориентации животного в пространстве и времени».

