 2015-06-26
2015-06-26 3894
3894Клеточная стенка образуется в результате развития срединной пластинки. Сразу после полного разделения ядра растительной клетки в телофазе митоза поперек делящейся клетки образуется фрагмопласт. Он состоит из множества уплощенных мембранных везикул — фрагмосом, содержащих компоненты клеточной стенки. В их построении участвует цитоскелет. Все полисахариды клеточной стенки, за исключением целлюлозы, синтезируются в аппарате Гольд-жи. Они упаковываются в везикулы, которые транспортируются к растущей
срединной пластинке и сливаются с ней. Срединная пластинка увеличивается по направлению к плазмалемме и соединяется с ней, разделяя две дочерние клетки. Наконец, вновь образующаяся клеточная стенка соединяется с уже существующей первичной клеточной стенкой.
Практически все «нецеллюлозные» компоненты клеточной стенки — полисахариды, структурные белки, широкий спектр ферментов — образуются в аппарате Гольджи и в его везикулах координированно направляются к клеточной стенке.
До сих пор не идентифицированы гены, кодирующие полисахаридсинтазы, участвующие в синтезе основных цепей «нецеллюлозных» полимеров. Идентифицированы гены нескольких фукозил- и галактозилтрансфераз, которые присоединяют отдельные сахара к главной цепочке.
Единственными полимерами, которые синтезируются с внешней стороны плазмалеммы, являются целлюлоза и каллоза. Целесообразность этого становится очевидной, если принять во внимание большую длину образующихся микрофибрилл целлюлозы и необходимость их филигранной укладки в клеточную стенку. Каллоза отличается от целлюлозы наличием β 1 → 3-D-глюкано-вых цепочек, которые могут образовывать спиральные дуплексы и триплексы. Каллоза образуется в нескольких типах клеток на определенных стадиях формирования клеточной стенки, например в прорастающей пыльцевой трубке или срединной пластинке делящихся клеток. Каллоза может также синтезироваться при стресс-реакциях или в ответе на грибную инфекцию.
Синтез целлюлозы катализируется мультимерными комплексами ферментов, расположенными на концах удлиняющихся микрофибрилл целлюлозы. Эти терминальные комплексы хорошо различимы под электронным микроскопом.
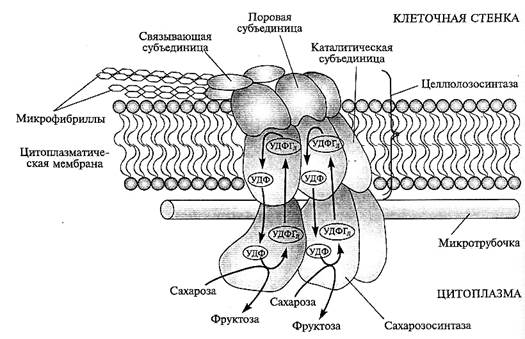
Рис. 1.30. Схема строения и работы целлюлозосинтазы
В некоторых морских водорослях терминальные комплексы синтеза целлюлозы расположены линейно; у всех покрытосеменных растений они формируют розеточные структуры. Терминальные комплексы появляются в мембране плазмалеммы в момент активации синтеза целлюлозы.
Исходным субстратом для целлюлозосинтазы является УДФ-глюкоза. Она образуется с помощью фермента сахарозосинтазы непосредственно из сахарозы. Ряд изоформ этого фермента находятся в плазматической мембране. Они ассоциированы с целлюлозосинтазой и могут поставлять УДФ-глюкозу непосредственно к ее каталитическому центру (рис. 1.30).
Относительно недавно были идентифицированы несколько растительных генов, кодирующих ферменты синтеза целлюлозы, в частности гены CesA, которые интенсивно экспрессируются в хлопковых волокнах во время активного синтеза целлюлозы вторичной клеточной стенки. Кодируемые этими генами полипептиды имеют восемь трансмембранных доменов и массу около 110 кДа. Открытие генов CesA дало возможность идентифицировать ряд других генов, кодирующих синтазы полисахаридов клеточной стенки.

