 2015-07-04
2015-07-04 6894
6894В нервной системе передача информации на длинные расстояния возможна благодаря тому, что потенциал действия распространяется вдоль аксона нервной клетки равномерно и без потери амплитуды. Аналогичным образом происходит распространение потенциалов действия по мембране мышечных волокон. Принцип распространения потенциала действия в этих структурах одинаков и связан с возникновением локальных токов. Мы уже знаем, что потенциал действия обусловлен возникновением входящего натриевого тока. При этом в соседних (неактивных) от места возникновения потенциала действия участках нервного или мышечного волокна возникают локальные выходящие токи, вызывающие перераспределение зарядов на мембране и деполяризацию в этих участках (рис. 25А). Как только деполяризация в неактивных участках достигает порогового уровня, в них открываются натриевые каналы и потенциал действия возникает по соседству
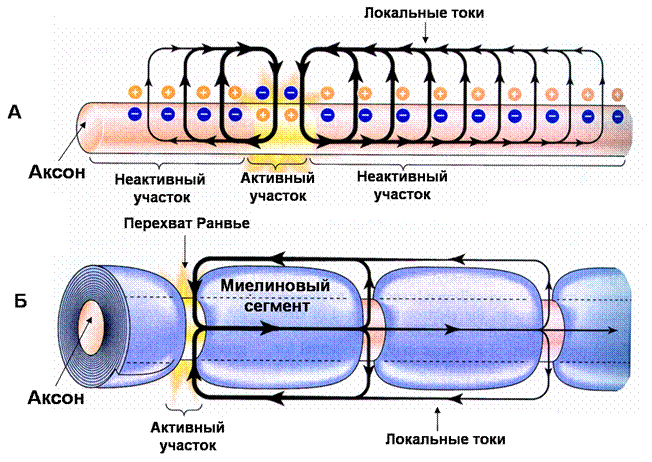
Рис 25. Проведение потенциала действия в немиелинизированных и миелинизированных нервных волокнах
(А) В участке мембраны немиелинизированного нервного волокна, охваченном потенциалом действия («активный участок»), возникают входящие локальные токи, которые возбуждают соседний, «неактивный» участок. Если мембрана не находится в рефрактерном состоянии, и деполяризация достаточна для достижения критического уровня деполяризации, то соседний участок становится активным и генерирует потенциал действия. Локальные токи затухают с расстоянием, поэтому в более отдаленных неактивных участках выходящий ток недостаточен для генерации потенциала действия.
(Б) В миелинизированном нервном волокне локальный ток может течь только через перехваты Ранвье, где нет миелина, и плотность натриевых каналов очень высока. Миелин обладает высоким сопротивлением, поэтому ток не течет в межперехватных участках. Активный участок представлен одиночным перехватом Ранвье. Поэтому потенциал действия распространяется скачкообразно от одного перехвата Ранвье к другому, а иногда перепрыгивает и через несколько миелиновых сегментов.
от первоначального места возникновения, и волна возбуждения продвигается дальше. Другими словами, проведение потенциала действия связано с его постоянным возникновением в соседних участках мембраны. Если взять изолированное нервное или мышечное волокно и вызвать в его середине потенциал действия, то он будет распространяться в обе стороны от места возникновения (рис. 25А). Однако при естественных условиях потенциалы действия распространяются по нервным волокнам только в одном направлении: от рецептора по дендриту к телу чувствительного нейрона и от тела нервной клетки по аксону к другой возбудимой клетке. Это связано с тем, что участки, расположенные сзади от продвигающегося потенциала действия находятся в состоянии рефрактерности, и локальные токи не способны вызвать в них возбуждение.
Скорость проведения потенциала действия зависит от того, насколько быстро и насколько далеко от активного участка происходит деполяризация мембраны до порогового уровня при протекании локальных токов. Это, в свою очередь, зависит от величины входящего тока, генерируемого в активном участке, и кабельных свойств волокна. Величина входящего тока зависит от плотности натриевых каналов в мембране, а кабельные свойства - от удельного сопротивления мембраны и аксоплазмы, а также от диаметра волокна. Оказалось, что чем толще нервное волокно, тем на большее расстояние будет распространяться деполяризация от активного участка, и тем больше скорость распространения потенциала действия. В гигантском аксоне кальмара, диаметр которого около 1 мм, скорость распространения равна 25 м/с, тогда как в некоторых нервных волокнах млекопитающих с диаметром меньше 2 микрометров скорость не превышает 1-2 м/с.
Для того, чтобы увеличить скорость распространения потенциала действия в относительно тонких нервных волокнах, природа создала на них специальную миелиновую оболочку (рис. 25Б). Так, в миелинизированных аксонах млекопитающих с диаметром всего 10-20 мкм скорость распространения равна 70-120 м/с. Миелиновую оболочку образуют глиальные
клетки: в периферической нервной системе - шванновские клетки, а в центральной – олигодендроциты. Мембрана глиальной клетки многократно наматывается на аксон и образует сегмент миелина длиной 1-1.5 мм. Между соседними сегментами миелина имеются короткие участки оголенной мембраны аксона, носящие название перехватов Ранвье. Сегменты миелина обладают изолирующими свойствами, а мембрана нервного волокна под ними практически не имеет проводимости и почти полностью лишена натриевых каналов. Области перехватов Ранвье, наоборот, имеют очень высокую плотность натриевых каналов. Поэтому локальные токи могут течь только от одного перехвата к другому, что позволяет деполяризовать мембрану на более длительные расстояния, вызывая в соседних перехватах Ранвье потенциалы действия. Такой способ проведения потенциала действия называется сальтаторным проведением и включает прыжки потенциала действия через миелиновые сегменты от перехвата к перехвату, что резко ускоряет продвижение потенциала действия по нервным волокнам.
По скорости проведения потенциала действия нервные волокна делятся на три группы (А, В, С). К группе А относятся миелинизированные нервные волокна диаметром от 20 до 2 мкм со скоростью проведения от 120 до 15 м/с. В зависимости от диаметра и скорости проведения волокна группы А часто подразделяются на 4 подгруппы (α, β, γ, δ). Эти волокна обеспечивают проведение возбуждения от рецепторов растяжения, прикосновения, боли, холода в нервную систему и от нервной системы к скелетным мышцам. К группе В относятся преганглионарные миелинизированные волокна с диаметром менее 3 мкм и скоростью проведения 3-15 м/с. И, наконец, к группе С – немиелинизированные нервные волокна с диаметром 0.3-1.3 мкм и скоростью проведения 0.5-2 м/с. Они образуют постганглионарные симпатические нервные волокна и волокна дорсальных корешков спинного мозга, ответственных за восприятие некоторых видов боли.

