 2015-07-04
2015-07-04 2793
2793Термин " синапс " был введен выдающимся английским физиологом Чарльзом Шеррингтоном в 1897 году после пионерских морфологических работ Рамон-и-Кахаля, в которых впервые были получены свидетельства о том, что нервная система состоит из изолированных нервных клеток – нейронов. Слово "синапс" переводится с греческого как соединение, связь. В настоящее время под синапсом понимают специфическое место контакта одной возбудимой клетки с другой, в котором происходит процесс передачи информации путем изменения потенциала мембраны. Основным в нервной системе является химический синапс, названный так потому, что в этом виде соединений в процессе передачи информации участвует химическое вещество - нейромедиатор (синонимы – нейротрансмиттер, медиатор, посредник). В таких синапсах одна клетка (пресинаптическая) обладает способностью синтезировать и выделять медиатор в окружающую среду (узкую щель между мембранами клеток – синаптическую щель), а другая (постсинаптическая) – воспринимать и отвечать на это вещество специфической реакцией в виде изменения своего мембранного потенциала (поляризацией) и возникновением постсинаптического потенциала. В зависимости от типа контактирующих клеток, различают два основных типа синапсов – нервно-мышечные и межнейронные.
Квантово-везикулярная гипотеза освобождения нейромедиатора. В 1953 году Бернард Катц с сотрудниками, исследуя нервно-мышечный синапс лягушки, с помощью микроэлектродов зарегистрировали новый класс постсинаптических потенциалов. Эти сигналы возникали случайно, в покое, имели очень маленькую амплитуду и были названы миниатюрными потенциалами концевой пластинки (МПКП). В ответ на раздражение двигательного нерва возникает высокоамплитудный постсинаптический потенциал, который получил название потенциала концевой пластинки (ПКП). ПКП от раздражения к раздражению варьируют по амплитуде, причем эти колебания кратны амплитуде МПКП. Было предположено, что медиатор в синапсе освобождается в виде мультимолекулярных порций – квантов. В покое случайное освобождение из нервного окончания отдельных порций вызывает появление на постсинаптической мембране МПКП, а в ответ на раздражение происходит синхронное освобождение нескольких десятков или сот квантов, и возникает ПКП. Электрофизиологическое определение показало, что квант нейромедиатора в нервно-мышечном синапсе состоит примерно из 10000 молекул ацетилхолина.
Использование метода электронной микроскопии для изучения ультраструктуры синапса позволило в 1954 году Де Робертсу и Беннету выявить в цитоплазме двигательного нервного окончания большое количество синаптических везикул диаметром около 50 нм. Поскольку везикулы имели одинаковые размеры и концентрировались у пресинаптической мембраны, было предположено, что квант медиатора находится в синаптической везикуле, а освобождение медиатора происходит путем выделения содержимого везикулы в синаптическую щель путем экзоцитоза.
Так возникла квантово-везикулярная гипотеза, согласно которой нейромедиатор выделяется в синапсе определенными порциями – квантами, аи кванты содержатся в специализированных органеллах – синаптических везикулах. В дальнейшем квантово-везикулярная гипотеза освобождения медиатора получила подтверждение на самых разнообразных объектах и в настоящее время считается общепризнанной.
За создание квантовой гипотезы секреции нейромедиатора и другие исследования, касающиеся механизмов синаптической передачи, в 1970 году Бернард Катц, совместно с Ульф фон Эйлером и Джулиусом Аксельродом были удостоены Нобелевской премии в области физиологии и медицины (рис. 27).

Рис. 27. Лауреаты Нобелевской премии 1970 года Сэр Бернард Катц, Ульф фон Эйлер, Джулиус Аксельрод
В настоящее время в центральной и периферической нервной системе обнаружено большое число медиаторов. Наиболее распространенные из них - ацетилхолин, глутамат, гамма-аминомасляная кислота (ГАМК), глицин, биогенные амины (норадреналин и дофамин), АТФ.
Передача возбуждения в химическом синапсе состоит из следующих принципиальных этапов (Рис. 28):
1) Деполяризация пресинаптической терминали во время потенциала действия;
2) Вход ионов Са2+ через потенциал-зависимые Са2+ каналы в пресинаптическую терминаль;
3) Освобождение нейромедиатора в синаптическую щель посредством экзоцитоза синаптических везикул;
4) Связывание нейромедиатора с постсинаптическими рецепторами;
5) Возникновение постсинаптического потенциала вследствие протекания ионных токов через каналы рецепторов.
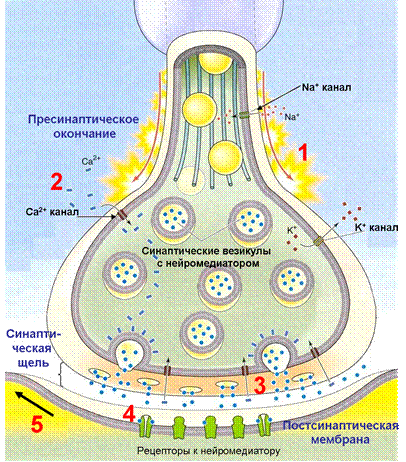
Рис. 28. Основные этапы химической синаптической передачи
Смотрите пояснения в тексте.
Далее мы подробно остановимся на механизмах возникновения постсинаптических потенциалов в нервно-мышечном синапсе и различных видах межнейронных синапсов.
В основе любого постсинаптического потенциала лежит активация ионотропных рецепторов постсинаптической мембраны к нейромедиатору, выделяемому пресинаптической клеткой.
Что представляет собой ионотропный рецептор?По своей сути он является лиганд-зависимым ионным каналом, активируемым при воздействии нейромедиатора с внеклеточной стороны.Основой всех ионотропных рецепторов является крупный белок, состоящий из пяти, реже четырех белковых субъединиц. Молекулярные массы субъединиц варьируют обычно в пределах от 40 до 70 кД. Первичная структура белков различных ионотропных рецепторов обнаруживает высокую степень гомологии. Субъединицы рецептора пронизывают толщу клеточной мембраны, образуя ионный канал. Участки полипептидных субъединиц, расположенные над поверхностью клетки, служат для узнавания и взаимодействия с нейромедиатором. Участки субъединиц, образующие собственно канал, характеризуются богатством гидрофобных неполярных аминокислотных остатков, обладающих высоким сродством к липидному (мембранному) окружению рецептора. Участки субъединиц, расположенные на внутренней поверхности мембраны, служат для взаимодействия с клеточными скелетными белками, ограничивающими подвижность рецептора, и являются мишенью для факторов, регулирующих активность рецептора в зависимости от ряда внутриклеточных процессов. Примером ионотропного рецептора служит никотиновый рецептор ацетилхолина (рис. 29).
Ионотропные рецепторы способны образовывать большое количество подтипов за счет различного сочетания субъединиц. Например, существует более 20 подтипов ГАМКА-рецепторов c различным комбинациям α-, β-, γ- и δ-субъединиц. В состоянии покоя каналы ионотропных рецепторов закрыты. При взаимодействии с медиатором происходит конформационная перестройка субъединиц рецепторов, и каналы открываются на несколько миллисекунд. После активации рецепторные макромолекулы теряют на некоторое время чувствительность к медиатору - наступает десенситизация.
Природа ионов, которые способен пропускать рецептор, определяется диаметром канала и характеристиками боковых радикалов аминокислотных остатков стенки канала. Никотиновые рецепторы ацетилхолина открывают путь ионам калия из клетки, а ионам натрия и кальция - внутрь клетки; глутаматные AMPA-рецепторы пропускают в основном одновалентные катионы, а NMDA-рецепторы – еще и ионы кальция; ГАМКА и глициновые рецепторы пропускают внутрь клетки ионы хлора.
Следует заметить, что помимо ионотропных, существуют и метаботропные рецепторы – трансмембранные белковые молекулы, передающие сигнал с мембраны клетки на G-белок и далее на белок-эффектор, катализирующий синтез того или иного вторичного посредника. Активация метаботропных рецепторов хотя и может опосредованно приводить к изменению проницаемости мембраны клетки для ионов, но не вызывает появления характерного постсинаптического потенциала. Некоторые нейромедиаторы (ацетилхолин, глутамат, ГАМК, АТФ) имеют как ионотропные, так и метаботропные рецепторы, тогда как другие (норадреналин, дофамин) - лишь метаботропные рецепторы.

