 2015-08-21
2015-08-21 7272
7272Наиболее крупный отдел растительного царства, насчитывающий более 350 семейств, 13 тыс. родов и до 240 тыс. видов. В настоящее время покрытосеменные распространены по всему земному шару — от тропиков до арктических и антарктических пустынь.
Автотрофные цветковые растения — важнейший активный компонент биосферы. От их жизнедеятельности зависит течение общебиосферных процессов обмена веществ и трансформации энергии, газовый состав атмосферы, климат, водный-режим суши, характер процессов почвообразования.
В наземных условиях цветковые обычно доминируют в растительном покрове, «каркас» большинства экосистем образуют именно они. Только в немногих сообществах и типах растительности они уступают господство по общей массе другим группам растений — в хвойных лесах голосеменным, в некоторых типах тундр й на сфагновых болотах — мохообразным, но и в таких случаях число видов покрытосеменных обычно превышает видовое разнообразие других отделов и подотделов растительного царства.
Понятно, что покрытосеменные определяют самую возможность существования большинства наземных животных. Они формируют среду обитания животного населения. Животные связаны с ними самыми разнообразными трофическими, консортивными и иными связями. Многие группы животных вообще могли возникнуть только тогда, когда Землю завоевали покрытосеменные и образуемые ими сообщества. Для многих членистоногих и позвоночных характерна сопряженная с цветковыми эволюция. Ясно, что и человек как биологический вид тоже не мог бы возникнуть и существовать, не будь на Земле покрытосеменных.
Покрытосеменные растения отличаются необычайным полиморфизмом, удивительной эволюционной пластичностью и могут существовать в самых разнообразных условиях обитания, вплоть до безводных пустынь, скал, солончаков, разбитых песков, шахтных терриконов, трещин в асфальте или бетоне. Это единственная группа среди высших растений, представители которой сумели вторично освоить морскую среду: в соленых водах морских мелководий вместе с водорослями растут десятки видов покрытосеменных.
Разнообразие размеров и жизненных форм цветковых растений, их приспособлений к конкретным условиям обитания поражает воображение. Самый крохотный их представитель, ряска волъфия (Wolffia arrhiza) едва достигает 1— 1,5 мм в диаметре и похожа на свободно плавающую зеленую водоросль, а попросту — на булавочную головку. С другой стороны, гигантские эвкалипты (виды рода Eucalyptus из семейства Myrtaceae) вырастают высотой до 100 м, а длина побегов тропических лиан — ротанговых пальм измеряется сотнями метром. Преобладают среди покрытосеменных автотрофные растения, но немало среди них полупаразитов, паразитов и сапротрофов, полностью утративших хлорофилл и способность к фотосинтезу. Эпифиты и эпифиллы (эпифиты, живущие на листьях других растений), деревья, кустарники, кустарнички, полукустарники и травы (многолетники и однолетники), плотоядные растения с их разнообразными приспособлениями для улавливания и переваривания мелких животных — это общеизвестные примеры. А тончайшие приспособления к опылению строго определенными агентами, к распространению плодов и семян, к перенесению неблагоприятных климатических периодов... Все это дает цветковым растениям возможность в полной мере реализовать свой эволюционно-адаптационный потенциал. Ни одна группа растений не могла выработать такого разнообразия приспособлений к определенным факторам среды.
ПРИЗНАКИ ПОКРЫТОСЕМЕННЫХ
По давней традиции характернейшим признаком покрытосеменных (греч. antos — цветок, phyton — растение) считают цветок — отсюда и название Anthophyta. Иногда утверждают даже, что понять и объяснить возникновение цветка — значит решить проблему происхождения покрытосеменных. Действительно, природа цветка — это одна из ключевых проблем морфологии и филогении, и споры по этому поводу, начавшиеся еще в XVIII в., продолжаются до сих пор. Однако даже общепринятого определения понятия «цветок» нет, и остаются неясными некоторые ключевые вопросы, касающиеся его природы.
Во-первых, цветки покрытосеменных необычайно разнообразны, и многие считают, что этот полиморфизм нельзя свести к какому-то единственному исходному прототипу. Во-вторых, хотя чаще всего типичный цветок определяют как так или иначе видоизмененный побег, строго говоря, не доказано, следует ли гомологизировать его с простым одноосным побегом или же с целой системой побегов. В-третьих, исторически сложилось так, что в поисках некоего архетипа за первичный примитивный цветок принимали совершенно разные образования. Так, сторонники эвантовой теории считают первичным крупный обоеполый цветок, опыляемый насекомыми, с большим и часто неопределенным числом свободных частей, и тогда все цветки однополые, олиго-мерные, голые во всех случаях рассматриваются как результат редукционной эволюции. Сторонники псевдантовой теории полагают, что эволюционные трансформации выражались в постепенном усложнении структуры цветка от однополых анемофильных с небольшим числом частей к обоеполым, полимерным, энтомофильным с хорошо развитым околоцветником. Есть и другие, нетрадиционные взгляды на происхождение цветка и его частей.
Истина, возможно, заключается в том, что нужно отказаться от поисков некоторого общего для всех покрытосеменных архетипа цветка и признать, что все современное разнообразие цветков сформировалось уже в пределах этой группы в ходе ее эволюции. Покрытосеменные сначала приобрели совокупность действительно важнейших для них общих признаков — в первую очередь уникальные особенности жизненного цикла, а уже потом выработали разные цветки. При этом какого-то одного генерального направления эволюции не существовало, и в каждой эволюционной ветви покрытосеменных были осуществлены различные морфогенетические решения.
Но при любых взглядах на цветок существенные его части — это андроцей, состоящий из одной, нескольких или многих тычинок (микроспорофиллов), и гинецей — из одного, нескольких или многих плодолистиков (мегаспорофиллов). Плодолистики могут быть свободными, и тогда каждый их них, срастаясь краями, образует пестик (один или несколько пестиков в цветке — по числу плодолистиков). При срастании нескольких плодолистиков формируется ценокарпный гинецей, пестик в нем всегда один.
Важнейшая часть пестика — завязь. Именно наличие завязи как замкнутой полости, в которой развиваются семязачатки (в отличие от голосеменных, у которых семязачатки расположены на поверхности открытых мегаспорофиллов), обычно считают важнейшим признаком покрытосеменных, откуда и название — Angiospermae (греч. aggeion — сосуд, вместилище, sperma — семя). Близко по смыслу и название, предложенное в свое время И.Н. Горожанкиным, — Gynoeciatae (пестичные). Полагают, что завязь обеспечивает лучшую защиту семяпочек. Но это, конечно, не защита от животных-пожирателей: стенка завязи большей частью слишком тонка и нежна, чтобы устоять перед мощными жвалами жуков или грызущим аппаратом гусениц бабочек. Вряд ли завязь способна надежно предохранить семязачатки и от неблагоприятных внешних воздействий, в том числе от колебаний температуры, хотя в некоторых случаях эта цель, по-видимому, достигается. Вероятно, смысл прежде всего в том, что в полости завязи семязачатки оказываются в условиях влажной камеры, оптимальных для их развития на определенном этапе. Кроме того, благодаря образованию завязи расширяются и обогащаются возможности распространения зачатков: из цветка формируется плод, и теперь при диссеминации разлетаться и расселяться могут уже не только отдельные семена, но и целые плоды, у которых возникают разнообразные приспособления к распространению семян самым широким кругом агентов.
Как и у некоторых голосеменных, у цветковых растений спермии доставлявются к яйцеклетке с помощью пыльцевой трубки — они «сифоногамны». Пыльцевая трубка обеспечивает высокую точность попадания спермиев в зародышевый мешок. Однако, коль скоро семязачатки скрыты в завязи и пыльца попасть непосредственно на микропиле не может, необходим какой-то специальный механизм, обеспечивающий улавливание пыльцы. Таким улавливающим аппаратом у покрытосеменных служит рыльце на верхушке столбика, или стилодия. Наличие рыльца — очень важная особенность. В свое время ван-Тигем даже предлагал называть покрытосеменные Stigmatae (рыльцевые, т.е. обладающие рыльцем).
У всех архегониальных растений четко прослеживается эволюционная тенденция к разноспоровости, сопровождающаяся редукцией полового поколения в жизненном цикле. Покрытосеменные представляют собой как бы завершающий этап на этом пути. Предельное сокращение процессов микро- и мегаспорогенеза и ускорение развития гаметофитов, связанное с их крайней редукцией, были, вероятно, важными эволюционными факторами. В результате женский гаметофит цветковых растений представлен зародышевым мешком, состоящим в типе всего из 7 клеток, а мужской гаметофит — пылинкой, в которой всего 2 клетки — сифоногенная и спермагенная, дающая 2 спермия; при развитии мужского гаметофита нет даже следов про-таллиальных клеток.
Одна из наиболее характерных особенностей покрытосеменных— двойное оплодотворение, при котором один из спермиев сливается с яйцеклеткой, образуя зиготу, а второй — с ядром центральной клетки зародышевого мешка, давая тем самым начало триплоидному эндоспер-м у. Это, по мнению ряда авторов, уникальный признак Angiospermae, аргумент в пользу их эволюционного единства и монофилетического происхождения. Часто высказываются соображения о возможности независимого и неоднократного возникновения процесса двойного оплодотворения в разных группах растений. Конечно, геометрически это можно себе представить, но с биологической и эволюционной точек зрения объяснение независимого в разных филах возникновения всей совокупности сложных механизмов, из которых складывается этот процесс, является дискуссионным.
Одно из следствий двойного оплодотворения — формирование своеобразной питательной ткани, полиплоидного эндосперма (в отличие от гаплоидного эндосперма голосеменных, представляющего собой женский гаметофит). В деталях биологическое значение полиплоидного эндосперма не объяснено, но, по-видимому, он обеспечивает более благоприятные условия для развития зародыша и поэтому мог сыграть важную эволюционную роль.
В вегетативной сфере подавляющее большинство покрытосеменных характеризуется наличием в ксилеме сосудов, тогда как почти у всех голосеменных древесина состоит только из трахеид. Лишь у немногих цветковых древесина гомоксильная, бессосудистая. Одни из них, как считают, сохранили этот анце-стральный признак, унаследовав его от предковых форм, а другие утратили сосуды в связи с существованием в особых экологических условиях, вроде водоемов, где подача воды по ксилеме большого значения не имеет. Вообще специализация тканей сыграла важную эволюционную роль. Паренхиматиза-ция позволила покрытосеменным выработать специальные запасающие ткани и разнообразные травянистые жизненные формы. В отличие от деревянистых растений у трав возможно сокращение онтогенеза и ускорение темпов эволюции; травянистые растения способны существовать в таких крайних условиях, в которых деревья и кустарники жить не могут.
ПРОИСХОЖДЕНИЕ ПОКРЫТОСЕМЕННЫХ
Ч. Дарвину происхождение покрытосеменных представлялось «отвратительной тайной». Сущность этой загадки, или тайны, состоит в том, что древнейшие достоверно определяемые ископаемые остатки цветковых растений встречаются лишь с начала мелового периода. В этих отложениях они очень немногочисленны и фрагментарны. Еще недавно считалось, что к середине мела покрытосеменные появляются в массе и выступают как «победители в борьбе за существование» с голосеменными и папоротникообразными. Сейчас палеоботаники смотрят надело несколько иначе и отодвигают начало расцвета покрытосеменных на более позднее время — конец мела.
Есть две альтернативные точки зрения. Одни исследователи склонны считать, что покрытосеменные возникли гораздо раньше мела — в триасе или даже палеозое. Первоначально небольшая, эта группа существовала, возможно, в таких условиях, которые не благоприятствовали захоронению их остатков, например, в горных районах или в пустынях. Позже, в мелу, Angiospermae, подготовленные к этому всей своей историей, дали мощную вспышку и сформировали в основном свое современное разнообразие.
Этому взгляду противоречат объективные данные палеоботаники. Многочисленные указания на нахождение остатков покрытосеменных в домел овых слоях, вплоть до обнаружения пыльцы в каменноугольном периоде, оказались несостоятельными: либо неверно был датирован сам возраст образцов, либо неправильна их идентификация, либо допущены какие-то методические неточности, либо, наконец, коллекционные образцы оказались утраченными, и проверить точность определения уже не представляется возможным...
Правда, косвенные указания на возможность существования покрытосеменных по крайней мере в юре есть. Это прежде всего некоторые формы насекомых, современные родственники которых настолько тесно связаны с цветковыми растениями, что допустить их независимое от последних существование трудно. Однако косвенные доказательства все же остаются косвенными. К тому же, зная об уже практически современных группах покрытосеменных в конце мела и допустив существование этой группы в юре, надо идти дальше и отодвинуть момент зарождения Angiospermae много глубже: иначе трудно объяснить, почему за десяток-другой миллионов лет они прошли основную эволюционную дифференциацию, а после этого, за 100—110 млн лет с конца мелового периода, мало изменились сколько-нибудь принципиально до наших дней. Как бы то ни было, достоверные остатки покрытосеменных в виде листьев, цветков, плодов, древесины и пыльцевых зерен известны начиная с мелового периода.
Согласно второй точке зрения, в предположении об очень раннем возникновении покрытосеменных нет надобности. Важно представлять себе эволюцию не просто как равномерный, не знающий скачков и спадов, векторизованный процесс. В ходе эволюции чередуются процессы арогенеза (того, что А.Н. Северцов применительно к морфологической эволюции животных называл ароморфозами) и аллогенеза (кладогенеза, телогенеза). В результате арогенеза — быстрого и важного по своим последствиям эволюционного скачка — группа резко и быстро вырывается на новый адаптационный уровень; она приобретает новые потенции и реализует их в процессе приспособительной, адаптивной эволюции на более высоком уровне. Если допустить, что возникновение покрытосеменных было результатом такого арогенетического процесса, то датировать это событие можно рубежом юры и мела. Это хорошо соответствует данным палеоботаники и позволяет без особых затруднений объяснить дальнейшую эволюцию и таксономическую дифференциацию цветковых растений.
В свое время пользовалась популярностью гипотеза о полярном происхождении покрытосеменных. Она привлекала специалистов возможностью сравнительно простого объяснения некоторых фактов удивительного сходства флоры Восточной Азии и приатлантической Северной Америки и вместе с тем показывала, какими путями цветковые могли проникнуть в Южное полушарие.
Однако эта гипотеза противоречит фактическому материалу, накопленному на сегодняшний день. Гораздо более вероятно, что родиной покрытосеменных были низкие широты, тропики и, возможно, субтропические районы. Какие именно — это служит предметом дискуссий. Одни исследователи высказываются за Юго-Восточную и Восточную Азию и Меланезию, т.е. области, прилегающие с запада к Тихому океану. В этом районе сосредоточены ареалы многих безусловно древних и считающихся примитивными таксонов покрытосеменных, а кроме того, сравнительно легко «прочитываются» вероятные пути миграций древнейших цветковых отсюда в другие области земного шара.
Согласно другой точке зрения, наиболее вероятно, что покрытосеменные возникли на древнем материке Южного полушария — Гондване и успели широко расселиться на его просторах до того, как Гондвана окончательно раскололась на современные материки к югу от экватора — Южную Америку, Африку, Австралию, Антарктиду. Эта гипотеза особенно привлекательна тем, что позволяет объяснить своеобразие и сходство флоры континентов и субконтинентов Южного полушария, разделенных сейчас практически непреодолимыми для цветковых растений океанскими пространствами. «Загадочные» ареалы некоторых таксонов, распространенных, например, в Юго-Восточной Азии и Южной Америке, также получают вполне естественное и логичное объяснение.
В качестве возможных предков покрытосеменных «испробованы» в разное время и с разной степенью обоснованности практически все группы высших растений, за исключением мохообразных, для которых характерен совершенно иной жизненный цикл с преобладанием гаметофита над спорофитом. Привлекали внимание и некоторые группы водорослей как реально существующие, так и чисто гипотетические (например, некая группа Chlorofloridae Б.М. Козо-Полянского, сочетающая черты зеленых и красных водорослей). В настоящее время наиболее распространено убеждение, что покрытосеменные все же представляют одну из ветвей того ствола, который объединяет все семенные растения, — следовательно, предками их могли быть какие-то голосеменные. Но дальше мнения расходятся.
Ряд исследователей считают, что вообще нельзя искать некоего общего предка для всех цветковых растений; разные их группы берут начало от различных предковых форм, иначе говоря, Angiospermae полифилетичны. В пользу этого взгляда свидетельствует прежде всего сам поразительный полиморфизм покрытосеменных, структурное разнообразие которых очень трудно свести к какой-то общей исходной модели, а также многообразие морфогенеза цветков в разных группах. Кроме того, данные палеоботаники говорят о том, что у разных голосеменных исторически вырабатывались отдельные черты сходства с покрытосеменными — у одних в вегетативной сфере, у других в строении репродуктивных органов. Общеизвестные примеры — обоеполые стробилы некоторых беннеттитовых, которые раньше рассматривались как прообраз цветка; купулы кейтониевых, в которых, как в завязи цветковых, укрыты семязачатки; микроспорофиллы с 4 пыльцевыми гнездами, как и тычинки покрытосеменных; сифоногамия у хвойных и оболочкосеменных и т.д. В пользу этих взглядов служит большое разнообразие цветков мелового периода — от однополых до обоеполых с синкарпным гинецеем.
Однако все эти черты сходства, по существу, представляют собой лишь реализацию отдельных попыток эволюции в разных эволюционных ветвях. Выявленные признаки сходства — это только некоторые внешние особенности, которые вполне могли возникнуть независимо и разными путями. Между тем таксон в филогенетической системе всегда характеризуется совокупностью признаков, и не обнаружена ни одна группа, которая связывала бы голосеменные и покрытосеменные, занимая промежуточное положение между ними не по каким-то отдельным признакам, а именно по их совокупности. Кроме того, по фрагментарным ископаемым остаткам практически невозможно судить, как был устроен у данного растения гаметофит, было ли ему свойственно двойное оплодотворение и т.п., т.е. насколько при внешнем сходстве оно обладало существенными чертами покрытосеменных.
Полифилетическое происхождение цветковых растений пока не подкрепляется прямыми фактическими данными. С точки же зрения строго эволюционной систематики вопрос стоит вообще в иной плоскости. Если мы считаем Angiospermae таксоном в том или ином ранге (отделом, подотделом, классом), то тем самым мы признаем его монофилетическое происхождение от одной общей предковой группы. Таксон филогенетической системы не может иметь полифилетическую природу. Если же мы признаем, что разные группы цветковых произошли от разных предков, то считать их единой группой оснований нет; тогда мы должны разбить покрытосеменные на столько таксонов равного ранга, сколько было предковых форм и эволюционных ветвей, приведших к формированию современных групп. Возможно, последующие детальные исследования разрешат, споры между монофилетистами и по-лифилетистами.
При всем удивительном полиморфизме цветковых растений общая уникальная совокупность их основных признаков все же дает основания считать их таксоном, имеющим общее происхождение, общего предка. Однако обычно внимание исследователей было сосредоточено прежде всего на объяснении возможного происхождения цветка. Считалось, что объяснив и поняв возникновение цветка, мы тем самым как бы объясняем и происхождение цветковых растений. Но, как уже говорилось, разные типы цветков могли, вероятно, формироваться независимо в различных группах.
Тем не менее именно из попыток понять происхождение наиболее типичного для покрытосеменных обоеполого цветка с так или иначе устроенным околоцветником родились основные гипотезы происхождения Angiospermae как таксона.
К началу XX в. сложились две противостоящие друг другу системы взглядов. Одна из них, связанная с именами австрийского ботаника Р. Веттштейна и классика немецкой ботаники А. Энглера, была основана на представлении о происхождении цветковых от эфедроподобных или гнетоподобньгх голосеменных предков. Р. Веттштейном была разработана оригинальная концепция происхождения цветка — так называемая псевдантовая теория, а точнее — идея о независимом возникновении частей цветка как органов «sui generis». Исходя из этой позиции, предполагалось, что первичными у покрытосеменных были раздельнополые опыляемые ветром цветки с небольшим и строго фиксированным числом частей, а дальнейшая их эволюция шла по линии «от простого к сложному».
Другая концепция восходит в конечном счете к типологическим построениям И.В. Гёте и А.П. Декандоля, т.е. к концу XVIII — началу XIX в., а в эпоху филогенетической систематики ознаменовались появлением стробиляр-н о й теории Арбера и Паркина. Согласно этой теории, наиболее близки к искомым предкам покрытосеменных мезозойские беннеттиты, а исходный тип цветка представляется сходным с тем, что наблюдается у многих современных многоплодниковых: обоеполый энтомофильный цветок с удлиненной осью, большим и неопределенным числом свободных частей. Дальнейшая эволюция цветка в пределах покрытосеменных имела редукционный характер.
Начиная с 30-х годов XX столетия все большей популярностью стали пользоваться взгляды сторонников телом ной теории, допускающих нетрадиционное истолкование происхождения частей цветка и, в частности, подвергающих сомнению возможность считать плодолистик органом листовой природы. Предложены многочисленные варианты, показывающие принципиальную возможность формирования разных типов цветков из различных пред-ковых структур, так или иначе сводимых к исходной системе стерильных и фертильных тел омов (гонофиллы Мелвилла, антокорм Мёзе и др.). Но все эти построения касаются более морфологической эволюции и не продвигают исследователей сколько-нибудь существенно в направлении поиска вероятной предковой группы цветковых.
Палеоботаника в последние десятилетия дала много нового материала о древнейших, ранне- и среднемеловых покрытосеменных. Эти данные позволяют сделать несколько заключений.
Во-первых, уже самые ранние из известных нам цветковых растений отличаются большим полиморфизмом. Очень разнообразны пыльцевые зерна — по форме, размерам, скульптуре поверхности, характеру поровости, числу и строению апертур. Они обнаруживают сходство с пыльцевыми зернами разных порядков и семейств современных растений. Листья меловых покрытосеменных также весьма разнообразны и напоминают современные фикусы, протейные, дубы, акации, аралию, подорожник и т.д.. Уже в мелу известны и сильно специализированные пельтатные (щитовидные) листья.

Листья некоторых меловых покрытосеменных
Большим полиморфизмом отличаются и органы размножения — цветки и плоды. У меловых цветковых известны как одиночные и относительно крупные цветки, так и мелкие, собранные в разнообразно устроенные соцветия с простым или двойным околоцветником, с апокарпным или ценокарпным гинецеем, с верхней или нижней завязью, с разным числом тычинок и плодолистиков. Наряду со вскрывающимися плодами типа листовок обнаружены и не-вскрывающиеся плоды. Вместе с деревянистыми формами в мелу определенно существовали и многочисленные травянистые покрытосеменные.
Во-вторых, если раньше часто утверждали, будто в мелу покрытосеменные были представлены уже современными таксонами, то сейчас, наоборот, большинство исследователей считают, что по крайней мере раннемеловые Angiospermae принадлежали к вымершим порядкам и семействам. Таким образом, предстоит еще выделить и описать эти группы и найти им места в общей схеме родословия цветковых. Но сам по себе этот факт очень важен как доказательство того, что покрытосеменные не оставались неизменными с момента рождения, но претерпели реальную эволюцию, в ходе которой к настоящему времени некоторые их представители исчезли с лица Земли.
Наконец, в-третьих, находки все новых и новых меловых цветковых растений нисколько не приблизили нас к решению вопроса об их вероятной предковой группе. Все эти ископаемые — это уже несомненные покрытосеменные, а форм, которые могли бы служить связующими звеньями между ними и их предками, так и не найдено. Возможно, конечно, что они существуют, но остаются неопознанными — надо постоянно иметь в виду трудность определения ископаемых остатков растений.
 | Соцветия, цветки и плоды ископаемых покрытосеменных |
В целом сейчас большинство специалистов склоняются к мысли, что предков Angiospermae нужно искать среди каких-то древних и малоспециализированных голосеменных. Вместе с тем не оставлена идея их генетической связи с беннеттитовыми, а многие склонны допускать полифилетическое и политоп-ное возникновение цветковых. Иногда даже утверждают (В.А. Красилов), что в ходе эволюции разных групп голосеменных имела место закономерная «ангио-спермизация». Но тогда покрытосемянность предстает как признак, характеризующий не таксон, а только определенный уровень организации репродуктивных структур.
Таким образом, уверенно ответить на вопрос о предках цветковых растений сейчас так же невозможно, как и во времена Ч. Дарвина. «Ископаемых остатков каких-либо растений, которые могли бы служить приемлемым связующим звеном между покрытосеменными и другими группами сосудистых растений, совершенно не известно. Нам приходится, следовательно, полагаться на дедукцию и умозаключения» (Г. Стеббинс).
КЛАСС ДВУДОЛЬНЫЕ (DICOTYLEDONEAE)
ПОРЯДОК ЦЕНТРОСЕМЕННЫЕ (Centrospermae)
СЕМЕЙСТВО ГВОЗДИЧНЫЕ (Caryophyllaceae)
Многолетние и однолетние травы, полукустарнички, часто со вздутыми в узлах стеблями, иногда с атипичным поликамбиальным утолщением. Листья супротивные, редко очередные, простые, часто не расчлененные на черешок и пластинку, с прилистниками или без них. Цветки в сложных цимозных соцветиях различного облика, реже одиночные, актиноморфные или слегка зигоморфные, как правило обоеполые, нередко (часто лишь функционально) раздельнополые. Околоцветник простой или двойной, цветок может иметь дополнительное покрывало из сближенных с ним прицветных листьев. Листочков простого околоцветника, чашелистиков и лепестков в типе 4—5, лишь в исключительных случаях их число большее и неопределенное. Лепестки часто хорошо дифференцированы на ноготок и отгиб, нередко глубокораздельный, иногда с адаксиальными придатками в основании. У части видов венчик полностью редуцирован. Тычинок в типе 10, в двух кругах. Тычинки любого круга могут редуцироваться. Гинецей из 2—5 плодолистиков, сросшихся в основании в верхнюю многогнездную или одногнездную завязь, с большим числом семязачатков. Плацентацию семязачатков гвоздичных часто называют свободной (центральной) колончатой, а их гинецей относят к лизикарпному типу. Плод — коробочка, иногда кожистая и невскрывающаяся («ягодообраз-ная»), или орех.
Около 80 родов и 2000 видов гвоздичных распространены по всему земному шару при явном предпочтении прохладных засушливых и холодных высокогорных или арктических областей. Поэтому наибольшее разнообразие гвоздичных присуще Северному полушарию, особенно Средиземноморью, а наименьшее — тропикам, где они встречаются в основном в горах.
 | Гвоздичные. Смолевка поникшая (Silene nutans): 1 — внешний вид; 2 — разрез через цветок. Гвоздика Фишера (Dianthus fischeri): 3 — внешний вид; 4 — диаграмма. Звездчатка дубравная (Stellaria nemorum): 5 — внешний вид; 6 — диаграмма цветка. Мокрицa (Stellaria media): 7 — разрез через цветок. Ясколка полевая (Cerastium arvense): 8 — разрез через цветок. Грыжник голый (Hernia-ria glabra): 9 — разрез через цветок. Дивала однолетняя (Scleranthus annuus): 10 — разрез через цветок. Смолка (Steris viscaria): 11 — диаграмма цветка |
Гвоздичные можно разделить на 3 подсемейства, рассматриваемые некоторыми авторами как особые семейства. Для подсемейства Silenoideae характерны сростнолистная чашечка и тенденция к образованию антофора — удлиненного участка цветоложа между чашечкой и венчиком. Плод — коробочка. Листья без прилистников. Ноготки лепестков часто с крыловидными выростами, а отгиб — с придатками в основании. Наиболее известны роды смолевка (Silene), зорька (Lychnis), мыльнянка (Saponaria). Корневища и корни представителей двух последних родов часто содержат много мылящих веществ — сапонинов. Для обширного рода гвоздика (Dianthus) характерны сближенные с цветком прицветники. Многие виды дали начало культурным сортам с яркими и крупными цветками. Цветки видов рода качим (Gypsophila) мелкие, собранные в сильно разветвленные соцветия. У пустынных и степных видов шаровидная надземная часть растения обрывается ветром по времени созревания плодов и, перекатываясь, распространяет семена. Интересен куколь обыкновенный (Agrostemma githago), встречающийся исключительно как полевой сорняк, в последнее время исчезающий. В подсемействе Alsinoideae чашелистики свободные, встречаются виды с односеменными невскрывающимися плодами. Наиболее известны виды родов звездчатка (Stellaria) и ясколка (Cerastium), приспособившиеся к очень разнообразным местообитаниям, нередко имеющие широкую экологическую амплитуду, становящиеся сорняками. Очень мелкие представители рода мшанка (Sagina) часто имеют редуцированный венчик, который полностью исчезает в роде дивала (Scleranthus). Для последнего подсемейства Paronychioideae в типе характерны безлепестные цветки, невскрываю -щиеся односеменные плоды, заключенные в чашевидный гипантий, часто прилистники. В отечественной флоре встречаются виды родов грыжник (Herniaria) и паронихия (Paronychia).
Двойной околоцветник гвоздичных, в отличие от аизооновых, возник за счет включения в цветок прицветных листьев, образовавших чашечку. Одно-покровность цветков и односемянность плодов связаны с редукционной эволюцией.
СЕМЕЙСТВО МАРЕВЫЕ, ИЛИ ЛЕБЕДОВЫЕ (Chenopodiaceae)
Многолетние и однолетние травы, реже полукустарники и кустарники или небольшие деревья с атипичным вторичным утолщением стеблей и корней. Иногда встречаются лианы. Листья очередные или, реже, супротивные, простые, без прилистников. Ксерофильные и галофильные маревые часто имеют глубоко уходящие в почву корневые системы, иногда утолщенные запасающие органы корневого или гипокотилярного происхождения. Им нередко присуща подушковидная форма роста, покрытые воском или опушенные, сочные цилиндрические, превращенные в колючки или редуцированные листья, сукку-лентные членистые побеги.
Цветки мелкие и невзрачные, обычно актщюморфные, обое- или раздельнополые. Они собраны в плотные клубочки, разветвленные по дихазиальному типу, которые, в свою очередь, часто объединены в сложные открытые соцветия колосовидного или метельчатого облика, редко цветки одиночные. Простой околоцветник из 3—5 свободных или слегка сросшихся листочков, иногда совсем редуцирован, часто при созревании плода не опадает и может сильно разрастаться.
Тычинки чаще всего в одинаковом числе с листочками околоцветника и противолежат им, или тычинок меньше — до одной. В основании тычинки объединены короткой трубочкой. Большей частью 2, реже 3—4(5) плодолистиков срослись в пестик с верхней одногнездной завязью. Единственный семязачаток прикрепляется к ее дну или боковой стенке. Плод односемянный, с пленчатым сухим перекарпием (мешочек), редко сочный, ягодовидный. Нередко он заключен в остающийся околоцветник и разросшиеся прицветники. Семена с периспермом.
 | Маревые. Марь белая (Chenopodium album): l — внешний вид; 2 —диаграмма цветка. Солерос (Salicornia europaea): 3 — диаграмма цветка. Сареазан (Halocnemum Strobilaceum): 4 — внешний вид. Солянка южная (Salsola australis): 5 — побег при плодах. Свекла (Beta vulgaris): 6 — цветок; 7 — его разрез |
Свыше 100 родов и около 1500 видов распространены по всему земному шару, особенно характерны для степных и пустынных районов, за пределами которых в основном являются рудеральными растениями и сорняками, часто злостными. Поэтому не случайно, что наиболее разнообразны маревые в сухих районах Австралии, в пампе Южной Америки, в Средиземноморье, Центральной и Юго-Западной Африке, в пустынях и полупустынях, Передней, Средней и Центральной Азии.
Построение системы семейства не завершено, в нем выделяют несколько триб, традиционно группируемых по форме и расположению зародыша в семени в 2 подсемейства. Для подсемейства Chenopodioideae (Cyclolobeae) характерен изогнутый зародыш, кольцом окружающий обильный перисперм. Космополитный, объединяющий до 200 видов род марь (Chenopodium) имеет обоеполые цветки. Марь белая (Ch. album) относится к числу наиболее распространенных злостных сорняков. Ее мощные побеги, как и у многих маревых, можно употреблять в пишу. Этот же вид, как и некоторые другие, некогда использовали и для получения съедобных семян, т.е. как зерновую культуру. Среди марей есть пряные, лекарственные, кормовые и красильные растения. Виды близкого по биологии рода лебеда (Atriplex) имеют раздельнополые цветки и остающиеся при плодах прицветники.
Среди них также встречаются злостные сорняки, лекарственные и красильные, а также кормовые и съедобные растения. Последние, однако, содержат вещества, вызывающие у животных светлой масти и светловолосых людей сильные аллергические реакции на солнце. Плоды шпината (Spinacia oleracea) полностью окружены одревесневающими и сросшимися в трубку прицветниками. Разнообразные сорта свеклы (Beta vulgaris) культивируют ради мясистых корнеплодов, используемых, в зависимости от сортовой группы, в пищу, на корм скоту или для получения сахара. Характерная для сорта форма корнеплода во многом связана со степенью участия корня и гипокотиля в его формировании. Известны и листовые сорта свеклы — мангольд. Представители рода солерос (Salicornia) широко распространенные галофиты с суккулентными членистыми побегами, листья которых сильно редуцированы.
Подсемейство Salsoloideae (Spirolobeae) объединяет виды со скудным или отсутствующим периспермом и сильно развитым, спирально закрученным в семени зародышем. Виды галофилъного рода солянка (Salsola) ранее широко использовали для получения поташа и соды. Некоторые из них содержат ценные для медицины алкалоиды. Представители рода саксаул (Haloxylon) — характерные для песчаных, часто засоленных пустынь, деревья высотой до 6 м. Закрепляя пески, они служат в этих районах практически единственным источником древесины, а также наряду с другими видами маревых практически единственным кормом для диких животных и скота.
ПОРЯДОК МАГНОЛИЕЦВЕТНЫЕ (Magnoliales)
СЕМЕЙСТВО МАГНОЛИЕВЫЕ (Magnoliaceae)
Деревья и кустарники с очередными цельными и цельнокрайними, реже лопастными листьями с прилистниками и крупными одиночными цветками. Цветки ациклические или спироциклические, обоеполые. Околоцветник простой, обычно венчиковидный, ярко окрашенный, из 6 или большего числа листочков, обычно в 3-членных кругах. Ось цветка сильно удлинена. Тычинок много, в неопределенном числе, часто мясистых, плоских с выраженным надсвязником. Стаминодии отсутствуют. Гинецей, как правило, апокарпный, из многих или нескольких плодолистиков, расположенных по спирали. Рыльца низбегающие. Семязачатки в различном числе вдоль брюшного шва, анатроп-ные, с 2 интегументами. Плод из нескольких листовок или орешков, часто шишкообразный. Семена с эндоспермом, часто на длинных семяножках. Характерна мясистая ярко окрашенная саркотеста. Пыльца 1-бороздная. Узлы многолакунные.
10 родов и 220 видов, главным образом в горных субтропических лесах Юго-Восточной Азии, Меланезии, Северной и Центральной Америки.
Несмотря на небольшой объем (всего 220 видов!), магнолиевые относятся к числу наиболее известных семейств. Это связано не только с высокими эстетическими качествами магнолий и тюльпанных деревьев, но и с их ролью в филогенетических построениях.
Крупнейший в семействе род магнолия (Magnolia) распространен по всему ареалу семейства, заходя даже в районы умеренного климата. На южных Курильских островах (Кунашир) встречается как редкость магнолия обратнояйцевидная (М. obovata) — единственный дикорастущий вид семейства в России. В Крыму и на Кавказе часто разводят магнолию крупноцветковую (М. grandiflora), родина которой — юго-восток США. У этой магнолии крупные кожистые вечнозеленые листья и белые душистые, с исключительно сильным ароматом цветы до 20 см в диаметре. Листочков околоцветника 6, иногда 9 или 12. После цветения каждый из многочисленных пестиков превращается в листовку, тесно прирастающую брюшным швом к удлиненной оси цветка. Благодаря этому вскрывание листовок происходит по спинному шву. Очень эффектны ярко-красные ягодообразные семена, выставляющиеся на длинных семяножках.
Особого упоминания заслуживают прилистники. Они срастаются в некоторое подобие капюшона, выполняя важную защитную функцию не только для пазушных, но и для верхушечных почек.
Не все магнолии вечнозеленые растения, некоторые, как упоминавшаяся М. obovata, — листопадные деревья. Многие из них цветут перед распусканием листьев. Голые, без листьев, деревья, усыпанные крупными белыми, палевыми или ярко-розовыми цветами, необыкновенно красивы. Высокие декоративные качества магнолий давно прочувствованы японскими и китайскими художниками. Они заняли прочное место в восточно-азиатском искусстве.
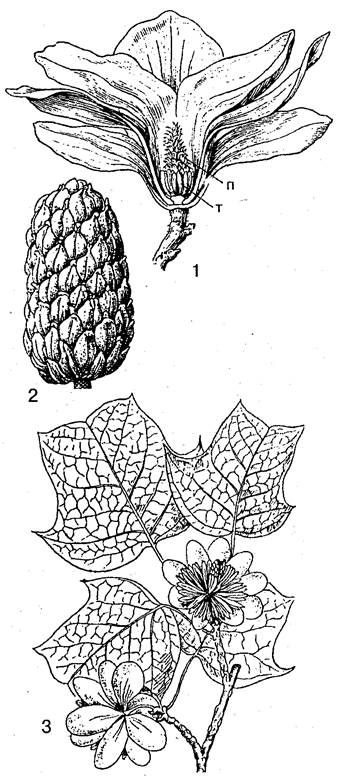 | Магнолиевые. Магнолия крупноцветковая (Magnolia grandiflora): l — продольный разрез через цветок (т — тычинки; п — пестики); 2 — плод из многих листовок, вскрывающихся по спинному шву. Тюльпанное дерево (Liriodendron tulipifera): 3 — цветущая ветвь |
Если у магнолий количество листочков околоцветника весьма различно и не всегда кратно трем, то у тюльпанного дерева (Liriodendron tulipifera) околоцветник строго 3-членный. Он состоит из 3 наружу отогнутых листочков и 6 вперед направленных и снабженных нектарниками, в 2 кругах. Впрочем, все листочки одинаково окрашенные, венчиковидные, а цветки схожи с тюльпанами (отсюда и название). Формулу такого околоцветника можно дать как К3 С3+3 или Кс3 GN3+3, отразив яркую окраску чашечки и наличие нектарников на лепестках. Как и у магнолий, в цветках тюльпанного дерева многочисленные тычинки и пестики сидят на удлиненной оси, однако плоды тюльпанного дерева своеобразны. Вместо листовок образуются орешки, причем верхняя стерильная часть каждого пестика преобразуется в крыло длиной несколько сантиметров. При созревании плода крыло все более и более оттопыривается от оси цветка, пока орешек не отрывается. При этом крыло действует как приспособление для полета и распространения семян.
Liriodendron tulipifera — мощное дерево высотой до 30 м, как и Magnolia grandiflora, родом с юго-востока США. Он разводится во многих местах Черноморского побережья Кавказа. Другой вид рода обитает в Юго-Восточной Азии. В отличие от магнолий листья у Liriodendron 4-лопастные. Из-за ряда своеобразных признаков (в дополнение к сказанному, отсутствие саркотесты) Liriodendron выделяют в особую трибу внутри семейства, противопоставляя всем остальным магнолиевым. Однако как и у всех магнолиевых, у него много-лакунные узлы и 1-бороздная пыльца. Обычный на нашем Дальнем Востоке лимонник (Schisandra chinensis), ранее относившийся к магнолиевым, сейчас выделен из последних, прежде всего по 1-лакунным узлам и 3—б-бороздной пыльце.
Семейство магнолиевых часто рассматривается как очень примитивное, и сторонниками «раналиевой» доктрины и стробилярной гипотезы цветка ставится вместе со следующим семейством в основание системы цветковых растений. С этих позиций к несомненно примитивным признакам семейства принадлежат слабая дифференциация околоцветника и нестабилизи-рованное число его членов, первичная полиандрия, листовидность и мясистость тычинок при выраженности надсвязников, апокарпный гинецей, низбегаю-щие рыльца, удлиненная ось цветка, 1-бороздная пыльца, 3-лакунные узлы, сосуды с лестничной перфорацией, наличие волокнистых трахеид. Нельзя полностью исключить и филогенетические связи предков магнолиевых с беннеттитами.
СЕМЕЙСТВО ДЕГЕНЕРИЕВЫЕ (Degeneriaceae)
Интересно в филогенетическом отношении.
Семейство представлено единственным видом Degeneria vitiensis (по имени немецкого ботаника Отто Дегенера, впервые собравшего растение; vitiensis — по острову Вити-Лёви в архипелаге Фиджи) на островах Фиджи. D. vitiensis — невысокое дерево с листьями, схожими с листьями магнолии, и небольшими одиночными цветками. Замечательные особенности Degeneria лежат в строении их тычинок и пестиков. Тычинки совершенно плоские, листоподобные со значительными стерильными участками и 3 (!) сосудистыми пучками. Ни связника, ни тычиночной нити здесь нет. Пыльники находятся на нижней стороне тычинки. Не менее своеобразен единственный пестик в цветке Degeneria. Завязь образована плодолистиком, который в молодости не срастается своими краями и остается, в отличие от громадного большинства покрытосеменных, незамкнутым. Рыльцевая поверхность соответствует краям плодолистика. Плоды Degeneria многосемянные, сухие, но без явных способов вскрывания. Есть у Degeneria и другие интересные особенности, как, например, непостоянное число семядоль (чаще всего 3).
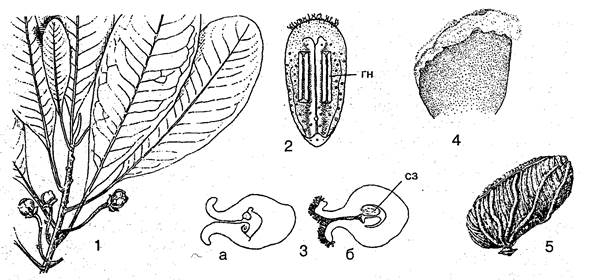
Дегенериевые.
Дегенрия (Degeneria vitiensis): 1 — цветущая ветвь; 2 — тычинка с нижней стороны (га — гнезда пыльника); 3 — завязь в поперечном разрезе на ранней (а) и более поздней (б) стадии развития (сз — семязачатки); 4 — пестик; 5 — плод
Дегенериевые, несомненно, сходны с магнолиевыми по строению околоцветника, из неопределенного числа членов, большому количеству тычинок, 3-лакунным узлам, 1-бороздной пыльце, а также по общему облику. Однако у дегенериевых отсутствуют весьма характерные для магнолиевых прилистники. Пестик у них один, и ось цветка почти плоская. Тычинки и плодолистики их примитивнее, чем у магнолиевых.
ПОРЯДОК ЛЮТИКОЦВЕТНЫЕ (Ranunculales)
СЕМЕЙСТВО ЛЮТИКОВЫЕ (Ranunculaceae)
Травы, редко полукустарники, невысокие кустарники или лианы с очередными цельными и рассеченными листьями без прилистников (реже листья супротивные). Цветки в разнообразных соцветиях, реже одиночные, энтомофильные, обоеполые, ациклические или циклические, актиноморфные, реже (колено Delphinieae) зигоморфные. Околоцветник разнообразный, простой или двойной, часто спорной морфологической природы, характерны стаминодии — нектарники и органы, промежуточные между стаминодиями и лепестками. Тычинок много или несколько, в неопределенном числе, редко 3—2 (1). Пестиков много, свободных, на удлиненной оси, реже 5—3—1. Завязь 1-гнездная верхняя. Рыльце часто низбегающее. Семязачатки в различном числе (вплоть до 1), вдоль брюшного шва, анатропные, с 2, реже 1 интегументами. Плоды из нескольких (до 1) листовок или орешков. Семена с эндоспермом и мелким зародышем. Семядоля часто 1. Первичных археспориальных клеток несколько. Нередко проводящие пучки — в нескольких кругах. Узлы 3- или многолакунные. Очень распространены алкалоиды.
40—50 родов и около 2000 видов в холодных, умеренных и субтропических районах Северного полушария, часто в горах, очень немногие виды (в основном из рода ломонос (Clematis) — в тропиках.
Лютиковые — обширное и исключительно разнообразное по строению цветков семейство. Наиболее примитивными считаются актиноморфные цветки, с простым околоцветником из неопределенного числа членов, большим числом тычинок и пестиков на сильно выпуклой оси. Примером может служить купальница европейская (Trollius europaeus) с пальчато-раздельными листьями и крупными ярко-желтыми цветами. Все части цветка купальницы расположены по спирали. Между листочками околоцветника и андроцеем располагаются несколько похожие на тычинки стаминодии. Весьма вероятно, что лепестки некоторых лютиковых могли возникнуть в процессе эволюции из таких стаминодиев. Стаминодии Т. europaeus — в то же время и нектарники. А у другого вида — купальницы азиатской (Т. asiaticus) стаминодии нектар не выделяют. Они вполне похожи на лепестки ланцетной формы. Их и можно считать лепестками, но тогда околоцветник Т. asiaticus двойной. Если же мы все же назовем их стаминодиями, околоцветник должен рассматриваться как простой. Подобная терминологическая двойственность обычна в литературе, посвященной лютиковым.
Цветки купальницы европейской имеют замечательную особенность — они цветут как бы в не совсем раскрытом состоянии (отсюда удачное французское название buton d'or — золотой бутон). Недавно открыли чрезвычайно специализированное опыление купальницы европейской мелкими двукрылыми. Этот факт лишний раз подтверждает, что виды, даже сохранившие примитивные признаки, прошли длительный путь эволюционных преобразований и приспособлены к своим экологическим нишам. Раньше купальница европейская принадлежала к обычным растениям средней полосы, теперь же благодаря истреблению на букеты встречается все реже и реже. В процессе эволюции цветки лютиковых подвергались, очевидно, следующим эволюционным преобразованиям: 1) цветки ациклические —> цветки гемициклические —> цветки циклические; 2) цветки с неопределенным числом членов -> цветки с определенным числом членов; 3) околоцветник простой —> околоцветник двойной (формирование двойного околоцветника в разных подсемействах происходило по-разному); 4) пестиков много -> пестик 1; 5) цветки актиноморфные -> цветки зигоморфные; 6) гинецей апокарпный -> гинецей синкарпный; 7) ось цветка удлиненная -> ось цветка почти плоская; 8) цветки неспециализированно энтомофильные -> цветки специализированно энтомофильные; 9) цветки неспециализированно энтомофильные -> цветки анемофильные. Как видим, лютиковые преобразовьшались по разным направлениям. Для многих родов характерно сочетание примитивных и подвинутых признаков.
Остановимся далее на самом крупном и широко известном роде лютик (Ranunculus), 400 видов которого распространены от арктических тундр до субтропиков. Для цветков лютика типична формула К5 CN5 A∞ G∞. Количество членов околоцветника здесь фиксированное, но лепестки имеют нектарные ямки, прикрытые специальной чешуйкой. Никакого разделения Лепестка на ноготок и пластинку (как, например, у крестоцветных и гвоздичных) здесь нет, однако детали строения самих нектарников довольно разнообразны. Лепестки-нектарники расположены по кругу, остальные части цветка — по спирали. Лютики опыляются широким комплексом насекомых из отрядов двукрылых и жуков, так что можно сказать, что их опыление очень неспециализированно, однако некоторые группы лютиков перешли к апомиксису, что является, безусловно, вторичным явлением.
Явная гетеробатмия характерна для родов аконит, или борец (Aconitum), и живокость (Delphinium), опыляемых исключительно перепончатокрылыми с весьма специализированными цветками. Напомним, что у однолетних живо-костей такие подвинутые признаки, как зигоморфные цветки со шпорцами, лепестки — нектарники, срастающиеся друг с другом, единственный пестик сочетается со спиральным расположением околоцветника и андроцея, множественностью тычинок и примитивностью пестика.
К ветроопылению перешел род василистник (Thalictram), большинство видов которого — высокие травы с дважды-, триждыперисторассеченными листьями и метельчатыми соцветиями из очень большого числа цветков. Околоцветник у василистников простой, мелкий и невзрачный. Тычинки, напротив, на длинных нитях, с ярко-желтыми или фиолетовыми пыльниками. Пестики, как и тычинки, в неопределенном числе, но их, как правило, меньше. Формула здесь может быть, например, такая: Рк4 A∞ G5-8.
 | Лютиковые. Лютик виноградолистный (Ranunculus cappadocicus): l — общий вид. Лютик едкий (Ranunculus cacris): 2 — лепесток с нектарной чешуйкой (н.ч); 3 — орешки. Лютик золотистый (Ranunculus auricomus): 4 — пестик в продольном разрезе. Лютик высокий (Ranunculus lingua): 5 — диаграмма цветка. Аквилегия альпийская (Aquilegia alpina): 6 — общий вид. Аквилегия обыкновенная (Aquilegia vulgaris): 7 — диаграмма цветка. Аконит (Aconitum napellus): 8 — общий вид; 9 — диаграмма цветка. Живокость (Delphinium datum): 10 — листовки. 3имовн к (Helleborus niger): 11 — нектарник |
Из сказанного ясно, насколько разнообразны цветки лютиковых. Однако они часто связаны переходными формами и имеют немало общего в деталях, например, в строении пестиков.
Для лютиковых очень характерны нектарники, в своем происхождении связанные с тычинками. Иногда они исключительно своеобразного, даже причудливого строения. Но у некоторых, например у василистников, их нет.
Почти все лютиковые — многолетние травы, есть среди них розеточные формы; для семейства очень характерны в различной степени расчлененные (но не сложные!) листья, без прилистников. Однако не так редко встречаются и цельные листья, например у чистяка (Ranunculus ficaria). Таким образом, узнавать лютиковые надо по совокупности признаков.
Многие лютиковые цветут очень рано, можно сказать, «из-под снега», в горах встречаются у тающих снежников. Очень немногие представители зацветают летом.
Лютиковые делят обычно на 2 подсемейства:
1. Зимовниковые (Helleboroideae). Плоды из листовок. К ним относятся купальница (Trollius), аконит (Aconitum), живокость (Delphinium) и др. Формирование двойного околоцветника происходило, по-видимому, за счет образования лепестков из стаминодиев, т.е. лепестки тычиночного происхождения.
2. Собственно лютиковые (Ranunculoideae). Плоды из орешков. К ним относятся лютик (Ranunculus), василистник (Thalictrum), ветреница (Anemone) и др. Формирование двойного околоцветника происходило, по-видимому, за счет образования чашелистиков из верхних вегетативных листьев, таким образом, и лепестки, и чашелистики имеют листовую природу.
Многие виды семейства декоративны. В садах часто можно встретить акониты, живокости, водосборы, или аквилегии, ветреницы. Особенно эффектны крупноцветные виды рода ломонос (Clematis), в том числе вьющиеся. Лютиковые дали ряд важнейших лекарственных растений. Здесь на первом месте адонис весенний (Adonis vernalis) — важнейшее средство при сердечно-сосудистой недостаточности. В настоящее время природные запасы этого степного растения сильно сокращены. Есть опыт его искусственного разведения. К классическим гомеопатическим средствам принадлежит «аконит», основанный на алкалоидах видов Aconitum. Лютиковые вообще богаты алкалоидами, в семействе много ядовитых растений. Некоторые горные среднеазиатские виды аконитов принадлежат к числу самых ядовитых растений.
Лютиковые — типичное семейство группы многоплодниковых (Polycarpicae), родственное магнолиевым и анноновым. В противоположность последним лютиковые — травянистое и более подвинутое семейство. В андроцее лютиковых, в частности, уже полностью отсутствуют «листовидные» тычинки, нет надсвязников, гораздо более специализирован околоцветник.
Долгое время к лютиковым причисляли род пион (Paeonia), который в настоящее время выделяется в особое семейство и даже порядок. У пионов, в отличие от лютиковых, вторичная, а не первичная полиандрия, своеобразный диск тычиночного происхождения и совершенно особый среди всех покрытосеменных эмбриогенез.
ПОРЯДОК МАКОЦВЕТНЫЕ (Papaverales)
СЕМЕЙСТВО МАКОВЫЕ (Papaveraceae)
Травы, реже полукустарники или кустарники, очень редко — небольшие деревья с очередными сильно рассеченными, реже цельными листьями без прилистников. Цветки одиночные или в различных соцветиях, обоеполые, более или менее циклические, 2—3-членные, правильные или зигоморфные (подсемейство Fumarioideae). Околоцветник двойной. Чашелистиков 2, очень рано опадающих. Лепестков 4, редко больше, в 2 кругах. Тычинок много, 8 или 2 с 3-раздельными тычиночными нитями. Завязь верхняя из 2 или многих (до 20) плодолистиков с 2—16 постенными плацентами и с большим числом анатропных или кампилотропных семязачатков с 2 интегументами. Столбики, или рыльца, сидячие. Плод — коробочка или стручок. Семена с эндоспермом и мелким зародышем, характерны млечники (не у всех). Содержат алкалоиды.
Более 45 родов и 700 видов в умеренных и субтропических районах главным образом Северного полушария.
По разнообразию цветков маковые напоминают лютиковые. В обоих семействах имеются актиноморфные и на первым взгляд совсем непохожие на них зигоморфные цветки. Однако маковые, как и лютиковые, представляют собой естественную группу, связанную общими признаками и переходными формами. Кстати, маковые вполне сопоставимы с лютиковыми по географии и родовому разнообразию (45—50), но едва ли не в 3 раза уступают последним по числу видов.
В целом для маковых характерен 2-членный тип цветка, особенно околоцветника, который соответствует формуле K2C2+2 как у актиноморфных, так и у зигоморфных цветков. Замечательный признак семейства — раннее опадание чашелистиков: их можно видеть лишь у бутонов; 2-членность у многих представителей проявляется в андроцее, однако у других, например у мака (Papaver), тычинок очень много. У видов последнего плодолистиков 5, 6, 12 и более, причем их число может варьировать даже внутри одного и того же вида. У большинства других родов плодолистиков 2, а плод представляет собой струч-ковидную коробочку. Такой плод можно видеть, например, у чистотела (Chelidonium majus), распространенного у нас возле жилья растения с желтыми цветками и оранжевым млечным соком). У мака плод — коробочка, вскрывающаяся дырочками. Формула цветка мака опийного (Papaver somniferum) K2C2+2AooG(5) –(20), формула цветка чистотела K2C2+2A12-22 G(2).
Мак, чистотел и другие роды с правильными цветками относятся к подсемейству собственно маковые (Papaveroideae). Представители с зигоморфными цветками объединяются в подсемейство дымянковые (Fumarioideaea). Познакомимся с ним на примере крупнейшего в семействе рода хохлатка (Corydalis), многочисленные виды которого (их всего 280) обитают в умеренной Евразии и Северной Америке, Многие хохлатки — геофиты с клубнями, как правило корневого происхождения, и эфемероиды — подснежники, как наша обычная в средней полосе хохлатка обыкновенная (С. solida), обитающая в широколиственных лесах. Это — красивоцветущее растение с кистями довольно крупных розово-фиолетовых цветов, интенсивно посещаемых шмелями. Цветки хохлатки резко зигоморфные прежде всего из-за вытянутого в шпорец одного из двух лепестков наружного круга. Если же сравнить диаграмму цветка хохлатки с диаграммой, например, аконита, то легко заметить, что тип зигоморфии здесь иной. Если у аконита плоскость, проведенная через ось соцветия, кроющий лист и центр цветка, совпадает с плоскостью зигоморфии, то у хохлатки она перпендикулярна последней. Подобный тип называется поперечной зигоморфией, и существует он только в семействе маковых. Венчик хохлатки имеет и еще ряд любопытных деталей, но, пожалуй, особенно интересен андроцей. Тычинок всего 2, каждая из них 3-раздельная и несет на верхушке по 3 пыльника. Средний пыльник нормальный, из двух половинок, боковые же фактически соответствуют половинкам. Предполагается, что предковая форма обладала правильными цветками и 4 тычинками, наподобие того, что мы видим у современного рода гипекоум (Hypecoum). Впоследствии, при переходе к зигоморфии в связи со специализацией опыления, помимо венчика сильно изменился и андроцей. Медианные тычинки расщепились, и их половинки срослись с боковыми тычинками. Формула цветка хохлатки может быть написана следующим образом: K2C2+2A2(1/2+1+1/2)G(2),. Плоды хохлатки — стручковид-ная коробочка. Семена обладают ариллусом и растаскиваются муравьями. Интересна у хохлатки и односемядольность, однако единственная семядоля возникает здесь в результате срастания (так называемая синкотилия), т.е. совершенно не так, как у однодольных.

Маковые.
Мак опийный (Papaver somniferum): 1 — общий вид; 2 — диаграмма. Макса-мосейкa (Papaver rhoeas): 3 — коробочка; 4 — она же в разрезе. Дицентра (Dicentra spectabilis): 5 — цветок (н.л — наружные лепестки; в.л — внутренние лепестки; т.н — тычиночные нити); 6 — схема строения пучка тычинок; 7 — диаграмма. Чистотел (Chelidonium majus): 8 — вскрывающийся плод. Гипекоум (Hypecoum): 9 — диаграмма цветка. Хохлатка (Corydalis): 10 — диаграмма цветка
К маковым относится одно из известнейших (хотя, скорее, печально известных) растений мак опийный (Papaver somniferam), в настоящее время распространенный только в культуре. Млечный сок, добываемый в основном из незрелых коробочек мака, содержит ряд алкалоидов, очень ценимых в медицине: морфин, наркотин, кодеин и др. Курение опиума, т.е. специально переработанного млечного сока, издавна было распространено, особенно в азиатских странах. В малых количествах опиум (в основном его составная часть морфин) вызывает приятное возбуждение, в больших — галлюцинации, нередко кончающиеся параличом нервной системы. Борьба правительств против ввоза и курения опиума иногда приводила к настоящим войнам, как, например, так называемая «опиумная война» между Англией и Китаем в XIX в., последовавшая после уничтожения китайцами более чем 20000 (!) ящиков опийного сырья. Интересно, что зрелые семена мака, употребляемые в кондитерской промышленности, совершенно не содержат морфина.
Выше была дана характеристика двух подсемейств маковых — собственно маковых (Papaveroideae) и дымянковых (Fumarioideae). Выделяют еще 3-е подсемейство гипекоумовые (Hypecooideae), куда относится упоминавшийся выше небольшой род гипекоум (Hypecoum). Поскольку из андроцея последнего выводят своеобразные тычинки — дымянковых и андроцей крестоцветных, Hypecooideae рассматривают часто в качестве исходной группы в семействе маковых. Тогда большое количество тычинок у мака и других родов, очевидно, вторичный признак. Однако этим соображениям противоречит то, что сам Hypecoum с его своеобразным венчиком, где лепестки внутреннего и наружного кругов сильно различаются, и однолетней жизненной формой создает впечатление производного рода. К тому же эмбриологические и палинологические признаки Hypecoum своеобразны.
Систематическое положение маковых не очевидно. Особенно спорно родство с нижеописываемыми семействами, часто объединяемыми с маковыми в один порядок Rhoeadales — Макоцветные. По-видимому, маковые имеют общих предков с лютиковыми. Интересны в этом отношении некоторые американские роды Papaveroideae, у которых встречаются такие признаки, как неопределенное количество лепестков и тычинок, коробочки, легко распадающиеся по числу плодолистиков, или 3-членный околоцветник К3С3+3. Однако они обладают млечниками, что не свойственно лютиковым.
ПОРЯДОК КАПЕРЦОВЫЕ (Capparales)
СЕМЕЙСТВО КАПЕРЦОВЫЕ (Capparaceae, или Capparidaceae)
Кустарники, невысокие деревья или травы с очередными простыми или пальчатыми листьями с прилистниками. Цветки одиночные или в кистевидных соцветиях, зигоморфные или актиноморфные. Прицветники отсутствуют. Характерен андрогинофор и/или гинофор, а также различные выросты оси цветка в виде диска или трубки, иногда имеется бокальчик осевого происхождения. Чашелистиков 4, в 2 кругах. Лепестков 4. Тычинок 4, 6 или более, иногда очень много. Завязь верхняя, из 2, реже нескольких плодолистиков, 1- или многогнездная, с постенной плацентацией и большим числом кампилотроп-ных семязачатков с 2 интегументами. Столбик с головчатым или 2-лопастным рыльцем. Плоды — стручковидная коробочка или ягодообразные. Семена без эндосперма.
Каперцовые включают 45 родов и 800 видов в сухих районах тропиков и субтропиков, немногие — на юге умеренной зоны. Обитают в саваннах, зарослях сухих кустарников и даже полупустынях и пустынях. Соответственно образу жизни они часто обладают побегами, превращенными в колючки, мелкими склерофи-цированными или сильно редуци

