 2015-10-16
2015-10-16 10526
10526Функции. Тело растения не может быть изолировано от внешней среды, так как оно существует в состоянии непрерывного с ней обмена веществ. Поэтому главной функцией эпидермы является регуляция газообмена и транспирации — испарения воды, регулируемого живыми тканями. В процессе эволюции эпидерма возникла очень давно, в самом начале приспособления высших растений к жизни на суше. Без нее немыслимо само существование высших наземных растений.
Необыкновенно тонкая и эффективная регуляция газообмена возможна благодаря замечательным структурным особенностям эпидермы.
Эпидерма, кроме того, имеет целый ряд других функций. Она препятствует проникновению внутрь растения болезнетворных организмов, защищает внутренние ткани от механических повреждений и придает органам большую прочность; через эпидерму могут выделяться наружу эфирные масла, вода, соли. Эпидерма может функционировать как всасывающая ткань. Наконец, эпидерма принимает участие в синтезе различных веществ, в восприятии раздражений, в движении листьев. Таким образом, эпидерма — многофункциональная ткань.
Строение. Клетки эпидермы плотно сомкнуты между собой (за исключением устьичных щелей) и имеют ряд других приспособлений, препятствующих потере воды. Однако важнейшая особенность эпидермы заключается в том, что она состоит из живых клеток, и пока эпидерма жива она выполняет свои регулирующие функции.
Эпидерма — сложная ткань, так как в ее состав входят морфологически различные клетки:
• основные клетки эпидермы;
• замыкающие и побочные клетки устьиц;
• трихомы, т.е. производные эпидермальных клеток в виде выростов, волосков.
Основные клетки эпидермы. Объемную форму этих клеток можно представить, рассматривая их с поверхности и на разрезах, проведенных перпендикулярно к поверхности органа. Боковые стенки (антиклинальные, т.е. направленные перпендикулярно поверхности органа) часто бывают извилистыми, что повышает прочность эпидермы. Наружные стенки обычно толще антиклинальных и внутренних.
Наружные стенки имеют сложное строение, выявленное методами световой и электронной микроскопии. Внутренний, наиболее мощный слой стенки составлен целлюлозой и пектиновыми веществами.
С наружной стороны вся эпидерма покрыта сплошным слоем кутикулы. Ранее представляли, что кутикула состоит из однородной массы кутина, однако выяснилось, что кутикула неоднородна. Внутри ее вкраплены включения растительных восков, причем как раз наличие воска в толще кутина понижает проницаемость для воды и газов. Воск может выделяться и на поверхность кутикулы, образуя чешуйки, палочки и другие структуры различной мощности и формы. На листьях капусты и плодах сливы этот воск образует сизый легко стирающийся налет.
На границе кутикулы и целлюлозного слоя находится прослойка пектиновых веществ. Эта прослойка непосредственно связана с пектиновыми веществами срединной пластинки, разделяющей соседние клетки эпидермы. С пектиновой прослойкой связана также система чрезвычайно тонких и разветвленных канальцев (прожилок), пронизывающих толщу кутикулы. Эти прожилки, обнаруженные с помощью электронного микроскопа, также содержат пектины. Думают, что вещества-предшественники, из которых образуются кутин и воски, вырабатываются живыми эпидермальными клетками, выделяются ими в пектиновые прослойки и прожилки и передвигаются по ним наружу, а потом превращаются в кутин и воски.
Описанное сложное строение эпидермальных оболочек не остается неизменным в течение жизни растения. Оно непрерывно меняется в зависимости от возраста органа и условий жизни растения, и эти изменения возможны только благодаря деятельности живых протопластов.
В протопластах основных клеток обнаруживаются тонкий постенный слой цитоплазмы с мелкими редкими хлоропластами (их часто называют лейкопластами) и ядро. В некоторых клетках эпидермы хорошо развиты эндоплазматический ретикулум и высокоактивный аппарат Гольджи, что указывает на энергичную синтетическую деятельность.
Иногда эпидерма состоит из нескольких слоев клеток. При образовании многослойной эпидермы клетки протодермы делятся один или несколько раз параллельно поверхности органа. Такая эпидерма отмечена преимущественно у тропических растений, живущих в условиях непостоянной обеспеченности водой (бегонии, пеперомии, фикуса). Можно думать, что многослойная эпидерма представляет собой водозапасающую ткань. Эту функцию может выполнять и однослойная эпидерма, если объем ее клеток велик, как, например, у зебрины, обычно изучаемой на лабораторных занятиях.
От многослойной эпидермы следует отличать гиподерму (греч. hypo — ниже, под), под которой понимают слой клеток, прилежащий к эпидерме изнутри, но возникающий независимо от нее из клеток основной меристемы и отличающийся по строению от глубже лежащих тканей. Гиподерма может выполнять различные функции. В одних случаях ее клетки имеют сильно утолщенные оболочки, которые усиливают изолирующие и механические свойства эпидермы; в других случаях в гиподерме накапливаются различные вещества (пигменты, танины и пр.).
Устьица. Среди основных клеток эпидермы размещены устьица в характерном для каждого вида порядке и числе. Отдельное устьице состоит из двух бобовидных замыкающих клеток, между которыми находится устьичная щель. Эта щель может то расширяться, то сужаться, регулируя транспирацию и газообмен. Клетки эпидермы, примыкающие к замыкающим клеткам, часто отличаются от остальных основных клеток, и тогда их называют побочными клетками устьица. Замыкающие клетки вместе с побочными образуют устъичный аппарат.
Механизм движения замыкающих клеток неодинаков у различных растений, но он всегда основан на том, что оболочки замыкающих клеток неравномерно утолщены, следовательно, форма клеток меняется при изменении их объема и неравномерном растяжении оболочек.
Изменение объема клеток устьичного аппарата происходит вследствие того, что в клетках меняется концентрация осмотически деятельных веществ. Предполагают, что клетки устьичного аппарата при раскрытии щели активно (т.е. против градиента концентрации и, следовательно, с затратой энергии) оттягивают от окружающих клеток ионы калия, что ведет к повышению осмотического давления в клетках и увеличению их объема за счет всасывания воды. Отток же ионов совершается пассивно, т.е. по градиенту концентрации. Тогда вода уходит из замыкающих клеток, их объем уменьшается и устьичная щель закрывается. В связи с потребностью в источнике энергии для активного транспорта ионов замыкающие клетки содержат многочисленные митохондрии в активном состоянии. В замыкающих клетках имеются хлоропласты с сильно развитыми тилакоидами. Они синтезируют углеводы, необходимые для активной деятельности митохондрий.
Работу замыкающих клеток можно продемонстрировать на модели. Два резиновых мешочка, подобные замыкающим клеткам, скреплены концами, а их поверхность оклеена лентами, чтобы воспрепятствовать растяжению в толщину и длину. При нагнетании воздуха мешочки изгибаются и открывают устьичную щель. Было обнаружено, что в стенках замыкающих клеток направление микрофибрилл целлюлозы сходно с расположением лент на модельных мешочках. Искривлению замыкающих клеток способствует то, что брюшные стенки (обращенные к щели) толще спинных стенок (обращенных в противоположную сторону) и, значит, растягиваются слабее.
Движение устьичных клеток имеет очень сложный характер, так как зависит от нескольких причин (света, температуры). Подробнее этот вопрос рассматривается в курсе физиологии растений.
Злаки, осоки и некоторые другие растения обладают устьицами особого типа. Их замыкающие клетки расширены на концах и сужены в средней части, а оболочки сильно утолщены посередине. При увеличении объема клеток тонкостенные участки раздуваются и раздвигают края щели. Интересно, что у злаков концевые участки обеих замыкающих клеток соединены сквозными отверстиями в оболочках, через которые протопласты соединены между собой. Вероятно, это помогает работе клеток как единой системы.
Число и распределение устьиц очень варьирует в зависимости от вида растения и условий жизни. У обычных растений луга, леса, а также у многих культурных растений число их колеблется от 100 до 700 на 1 мм2 поверхности листа. Такое колоссальное число находит объяснение с чисто физической точки зрения. Если сосуды, наполненные жидкостью, закрыть пластинками с отверстиями и измерить скорость испарения, то окажется, что при одной и той же суммарной площади отверстий скорость испарения тем выше, чем больше число отверстий (закон Дальтона).
Измерения показали, что если устьица полностью открыты, то транспирация идет с такой же скоростью, как если бы эпидермы вовсе не было. При закрытых же устьицах транспирация резко снижается и фактически может идти только через кутикулу. Таким образом, эпидерма эффективно регулирует газообмен и транспирацию в широком диапазоне.
Трихомы. Клетки эпидермы образуют наружные выросты, которые объединяют под названием трихомов (греч. trichos — волос). Трихомы отличаются крайним разнообразием, оставаясь вместе с тем вполне устойчивыми и типичными для определенных видов, родов и даже семейств. Поэтому признаки трихомов широко используются в систематике растений в качестве диагностических.
Кроющие трихомы:
1 - 3- простые одноклеточные; 4 - простой многоклеточный; 5 - ветвистый многоклеточный; 6- простой двурогий; 7,8 — звездчатый (в плане и на поперечном разрезе листа)
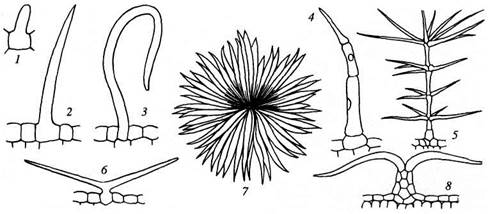
Трихомы бывают железистые и кроющие. Железистые трихомы образуют вещества, которые рассматриваются как выделения (экскреты или секреты). Они будут рассмотрены как выделительные ткани.
Кроющие трихомы имеют вид простых, разветвленных или звездчатых волосков, одно- или многоклеточных. Кроющие трихомы могут длительное время оставаться живыми, но чаще они быстро отмирают и заполняются воздухом.
Образованный волосками шерстистый, войлочный или иной покров отражает часть солнечных лучей и уменьшает нагрев, создает затишное пространство около эпидермы, что в совокупности снижает транспирацию. Иногда волоски образуют густой покров только там, где расположены устьица, например на нижней стороне листьев у мать-и-мачехи.
Однако живые волоски, увеличивая общую испаряющую поверхность, могут способствовать и повышению транспирации, что полезно растениям, живущим в условиях, когда транспирация затруднена.
От трихомов, возникающих только из эпидермальных клеток, следует отличать эмергенцы (лат. emergere — выдаваться), в образовании которых принимают участие и более глубоко расположенные ткани.

