2015-10-13
2015-10-13 492
4921. Дорсальные (задние) столбы спинного мозга представляют собой мощную систему волокон различного происхождения (рис. 6.21). Задние столбы у наземных млекопитающих в эволюции формируются в связи с развитием конечностей, когда возникает необходимость в тонком анализе информации от суставов и кожи, без которого невозможны ходьба, бег, прыжки, удержание равновесия и положения тела в пространстве. Прогрессивное развитие задних столбов в эволюции позвоночных животных тесно связано с развитием соматосенсорной коры больших полушарий.
Волокна задних корешков, войдя в спинной мозг, делятся на две веточки: длинную (восходящую) и короткую (нисходящую). Ядер задних столбов (нежного и клиновидного) в продолговатом мозге достигают лишь около четверти миелинизированных волокон, остальные аксоны оканчиваются на интернейронах спинного мозга.
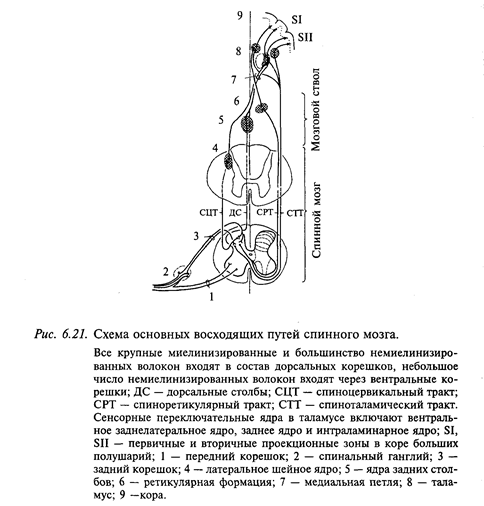
Аксоны нейронов ядер задних столбов (нежного и клиновидного) перекрещиваются и образуют медиальные лемниски, которые направляются к ядрам вентробазального таламуса. Волокна, составляющие задние столбы в спинном мозге, постепенно истончаются, так как по ходу отдают большое число коллатералей к интернейронам спинного мозга, поэтому скорость проведения импульсов снижается с 90-120 м/с - в месте вхождения в спинной мозг до 20-40 м/с на уровне шейных верхних сегментов спинного мозга.
В системе волокон задних столбов проходят афференты от многих типов рецепторов, в частности у животных - от рецепторов волосяных луковиц, суставов, подушечек лап, основания когтей и других образований. Мышечные афференты от конечностей также восходят в составе задних столбов и заканчиваются на нейронах ядер продолговатого мозга, а также на нейронах дорсального рога шейных верхних сегментов спинного мозга. Аксоны этих нейронов 2-го порядка перекрещиваются и присоединяются к волокнам медиального лемниска, которые, переключаясь через ядра вентробазального комплекса, проецируются на кору больших полушарий на границе сенсорной и моторной зон (поле За, по Бродману). Значительная часть афферентных волокон от мышечных и сухожильных рецепторов не входит в состав задних столбов. Эти волокна, начиная с поясничных сегментов спинного мозга и выше, переключаются на нейронах кларковых столбов, аксоны которых образуют мощные сенсорные тракты к мозжечку.
Суставные сумки и поверхности суставов имеют многообразные рецепторы, которые объединяются под общим названием суставные рецепторы. Согласно современным данным, в суставном нерве выделяют два типа ответов: быстро адаптирующиеся и медленно адаптирующиеся, последние более многочисленны. Соответственно этому выделяют два типа рецепторов. Медленно адаптирующиеся рецепторы, являясь наиболее типичными суставными рецепторами, сообщают о положении костей, образующих данный сустав. В суставных нервах обнаруживаются волокна, в которых разряды возникают как при сгибании, так и при разгибании сустава. Обнаружены также волокна, разряд которых в зависимости от сгибания или разгибания сустава, учащается или урежается (дирекционно-чувствительные ответы). Одно и то же движение сустава может вызвать урежение импульсации в одном афферентном волокне и увеличение ее в другом волокне. Информация от суставных рецепторов в супраспинальные центры проводится по системе волокон задних столбов.
Волокна задних столбов проводят также афферентные сигналы от внутренних органов. Импульсы от тазового нерва достигают надсегментарных структур не только по задним канатикам (задним столбам), но также и по вентролатеральным канатикам, в составе спиноталамических трактов спинного мозга. Скорость проведения импульсов от внутренних органов в задних столбах, например, кошки составляет 35-75 м/с, в вентральных-21-38 м/с и латеральных-17-32 м/с.
2. Спиноцервикальный тракт образован аксонами нейронов, тела которых находятся в основании серого вещества дорсального рога спинного мозга (см. рис. 6.21). Далее волокна тракта проходят ипсилатерально в составе латерального канатика и оканчиваются на нейронах шейного латерального ядра, локализованного в верхних сегментах шейного отдела серого вещества задних рогов спинного мозга. На этом же уровне аксоны нейронов этого ядра перекрещиваются и в составе специальной порции медиального лемниска направляются в вентробазальный комплекс таламических ядер.
Известно, что шейное латеральное ядро у человека представлено рассеянными клетками в шейных сегментах спинного мозга. У кошки, собаки, низших обезьян это ядро развито очень хорошо. На нейронах спиноцервикального тракта конвергирует возбуждение от низкопороговых кожных рецепторов, в том числе от рецепторов волосяных луковиц. Ряд нейронов возбуждается при пощипывании или давлении на кожу; размер их рецептивных полей может быть большим и занимать даже всю ипсилатеральную половину тела. Несмотря на большее количество переключений (спинальный уровень, шейное латеральное ядро, вентробазальный таламус, кора больших полушарий), возбуждение по спиноцервикальному тракту прибывает в кору больших полушарий на 2-5 мс быстрее, чем по волокнам дорсальных столбов. Это объясняется тем, что скорость проведения по волокнам задних столбов меньше, чем по волокнам спиноцервикального тракта.
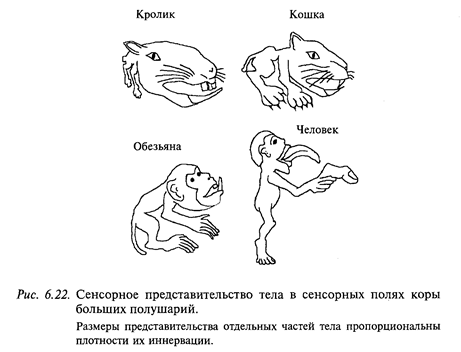
У млекопитающих (в том числе и у человека) по дорсальным столбам и спиноцервикальному тракту (у человека преимущественно по дорсальным столбам) в постцентральной коре больших полушарий организуется проекция всего тела (рис. 6.22). При этом соблюдается следующий принцип: чем выше плотность рецепторов в коже, покрывающей соответствующую часть тела, тем большей площадью эта проекция представлена в коре больших полушарий. Обратите внимание на то, что руки, особенно большой палец, оральная область, в том числе язык, голосовые связки и пр. представлены в коре мозга человека особенно подробно.
3. Спиноталамический тракт составляют самые тонкие миелинизированные афференты (диаметром до 6 мкм), а также немиелинизированные волокна задних корешков. Они входят в серое вещество спинного мозга и заканчиваются на нейронах основания серого вещества заднего рога. В сером веществе спинного мозга аксоны части этих нейронов перекрещиваются, а другая часть идет ипсилатерально, образуя спиноталамический тракт. Таким образом, спиноталамический тракт проецируется в ядра таламуса (вентральное заднелатеральное, парафасцикулярное и центральное латеральное ядра) билатерально. Часть нервных волокон тракта переключается на нейронах мозгового ствола, аксоны части этих нейронов переходят на контралатеральную сторону и также направляются в таламус.
По волокнам спиноталамического тракта передается информация о боли, давлении и температуре. Нейроны этого тракта имеют обширные кожные рецептивные поля, располагающиеся у некоторых нейронов билатерально, покрывая обе задние или обе передние конечности, а иногда даже всю поверхность тела. Нейроны, активируемые при раздражении одной конечности, как правило, тормозятся при стимуляции другой, тогда как нейроны с ограниченными кожными рецептивными полями часто тормозятся с окружающей кожной поверхности. В настоящее время считают, что этот тракт проводит генерализованные ощущения и по нему передается информация только о качестве раздражителя и его грубой локализации.
Таким образом, в таламус проецируются две системы: задние столбы (у ряда видов млекопитающих также спиноцервикальная система) и спиноталамический тракт. Эти две системы различаются функционально: первая проводит информацию о месте и модальности раздражителя (протопатическая чувствительность), тогда как вторая обеспечивает генерализованные формы ощущения (эпикритическая чувствительность).