 2015-10-22
2015-10-22 13490
13490Численность популяции = Численность в начальный момент времени + (рождающие особи + иммиграция) – (гибнущие особи + эмиграция).
Скорость изменения численности = (рождаемость + скорость иммиграции) – (смертность + скорость эмиграции).
Аналогично, формула для смертности:
1. Скорость изменения численности популяции.
Скорость изменения численности популяции можно определить путем деления величины изменения количества особей на период времени, за который оно произошло.
Средние скорости изменения популяций принято выражать в виде
V = DN / Dt,
где N - размер популяции (или другой показатель), t - время.
Например, предположим, что популяция из 100 представителей в некотором объеме воды за час увеличилась до 150 особей. Тогда DN = 50, Dt = (150 - 100) * 1 = 50 особей в час.
2. Рождаемость - способность популяции к увеличению численности за счет размножения. Различают максимальную и экологическую рождаемость.
· Максимальная, или физиологическая рождаемость - появление теоретически максимально возможного количества новых особей в идеальных условиях, то есть при отсутствии лимитирующих факторов. Этот показатель - постоянная величина для данной популяции.
· Экологическая, или реализуемая, рождаемость обозначает увеличение популяции при фактических, или специфических, условиях среды. Она зависит от состава, размера популяции и фактических условий среды.
Рождаемость обычно выражают:
· в виде скорости, определяемой путем деления числа вновь образовавшихся особей за определенный промежуток времени (dNn / dt) - абсолютная рождаемость)

· числом новых особей в единицу времени на единицу популяции (специфическая, удельная рождаемость).

где DNn – прирост численности, N – размер популяции или только части, способной к размножению (число особей в начальный момент времени).
Удельная рождаемость необходима для сравнения популяций с разной численностью.
| Удельная рождаемость выражается в %, или количеством особей на самку. Например, для высших организмов рождаемость выражают в расчете на одну самку. |
Рождаемость может быть нулевой или положительной, но никогда - отрицательной.
· Максимальная рождаемость - это теоретический верхний предел, который популяция могла бы достичь в идеальных условиях.
Несмотря на трудности при практическом определении этого показателя, он представляет интерес по следующим причинам.
1. Максимальная рождаемость служит критерием для сопоставления с реальной рождаемостью. Например, показатель рождаемости в популяции птиц 4 птенца в год будет иметь реальный смысл, если известен верхний предел, до которого она могла бы увеличиваться в менее лимитирующих условиях.
2. Так как максимальная рождаемость - величина постоянная для данной популяции, этот показатель можно использовать для математических расчетов и прогнозирования скорости роста популяции.
Максимальная рождаемость как характеристика популяции ценна тем, что с ней, как с константой, можно сравнивать различные наблюдаемые величины рождаемости.
Следует отметить, что понятие "максимальная рождаемость" относится к популяции, а не к изолированным особям.
За меру рождаемости следует принимать среднюю, а не индивидуальную наибольшую или наименьшую репродукционную способность.
Наиболее корректно определение максимальной рождаемости не только при отсутствии лимитирующих физических факторов, но и при оптимальных размерах популяции.
При сравнении рождаемости в популяциях разных видов возникают трудности, связанные с несопоставимостью жизненных циклов.
Поэтому при изучении рождаемости необходимо учитывать цикл развития организмов, особенно когда сравниваются различные популяции. В этом случае необходимо быть уверенным, что сравнение вообще возможно.
3. Смертность.
Этот показатель характеризует гибель особей популяций за определенный период времени. Так же, как и в случае с рождаемостью, смертность можно выразить числом особей, погибших за определенный период.
Различают:
· специфическую смертность - число смертей по отношению к числу особей, составляющих популяцию.
· экологическую или реализуемую, смертность - гибель особей в конкретных условиях среды (величина непостоянная, изменяется в зависимости от состояния природной среды и состояния популяции).
Смертность выражают числом особей, погибших за определенное время, выраженным в процентах от начальной численности популяции.

Как и в случае с рождаемостью, необходимо принять минимальную смертность за теоретическую константу.
Существует некая минимальная величина, характеризующая гибель особей в идеальных условиях, когда на популяцию не воздействуют лимитирующие факторы. В этих условиях максимальная продолжительность жизни особей равна их физиологической жизни, которая в среднем выше экологической продолжительности жизни.
Даже в идеальных условиях в любой популяции происходит смерть от старости, то есть имеет место какая-то минимальная смертность, определяемая физиологической продолжительностью жизни особей.
В природе средняя продолжительность жизни особей намного меньше, чем потенциальная, и фактическая величина смертности выше минимальной.
Смертность, как и рождаемость, особенно у высших организмов, сильно варьируется с возрастом, поэтому при изучении смертности организмов популяцию целесообразно разделить на возрастные группы.
таблицы выживания
Целостное представление о смертности в популяциях дают таблицы выживания (один из методов статистического анализа популяций), в которых приводится возрастная структура по продолжительности жизни особей.
Различают следующие типы таблиц выживания:
1. Когортные (динамические). Прослеживается история существования только одной когорты особей (от момента появления до гибели всех ее особей). При этом когортой мы называем группу особей родившихся в течение определенного промежутка времени. Часто применяется для изучения популяций однолетних организмов (насекомых).
2. Статические таблицы выживания. Вместо наблюдения за отдельной когортой ведут наблюдение за возрастной группой. При этом расчитывается смертность во всех возрастных группах.
Сложность построения таких таблиц заключается в невозможности реконструкции численности особей разных возрастных групп, с момента их рождения.
Они могут содержать следующие графы:
· возраст (временные промежутки, стадии развития (для личинок));
· число особей доживших до начала возрастного интервала, nx;
· доля особей доживших до возраста x, lx,;
· число особей, погибших в каждом возрастном интервале (x) – (x+1), dx;
· смертность в течение интервала x, qx;
nx+1=nx – dx
qx=dx / nx
lx=nx / n0
n0 – число особей начального возраста.
Таблица выживания усоногого ракообразного Balanus grandula (по Connell, 1970)
| Возраст, годы х | Число выживших к моменту х, nx | доля особей доживших до возраста x, lx | число особей, погибших в каждом возрастном интервале (x) – (x+1), dx; | смертность в течение интервала x, qx; |
| 0.56 | ||||
| 0.43 | 0.45 | |||
| 0.24 | 0.41 | |||
| 0.14 | 4.5 | 0.22 | ||
| 0.10 | 4.5 | 0.29 | ||
| 0.07 | 4.5 | 0.40 |
Таблица выживания популяции рисового долгоносика
| Середина возрастного интервала, недели | Удельное выживание, Lx | Удельная рождаемость, число потомков на 1-у самку возраста Х, mx Lx*mx | |
| 4,5 | 0,87 | 20,0 | 17,4 |
| 5,5 | 0,83 | 23,0 | 19,0 |
| 6,5 | 0,81 | 15,0 | 12,1 |
| 7,5 | 0,80 | 12,5 | 10,0 |
| 8,5 | 0,79 | 14,0 | 9,8 |
| 9,5 | 0,77 | 12,5 | 9,2 |
| R0=77,5 |
Зная показатели удельной рождаемости (mx) по таблице выживания можно определить
· чистую скорость размножения (воспроизводства), что составляет число потомков на каждую самку для всей популяции.
 (в данном примере = 77,5 особей)
(в данном примере = 77,5 особей)
В стабильных условиях в природе величина R приблизительно = 1.
| R0показывает во сколько раз может увеличится численность популяции за одно поколение. (То есть сколько особей может появиться в будущем на 1-у особь сейчас) |
Если построить график зависимости lx от Х, то получим
· кривую выживания (п о числу доживших до данного возраста организмов популяции).

Выделяют три типа кривых выживания:
· сильновыпуклые кривые (I) - характерны для видов организмов в популяциях, смертность которых почти до конца жизненного цикла остается низкой. Такой тип кривых выживания характерен для многих видов крупных животных, в том числе и для человека;
· сильновогнутые кривые (IV) - характерны для видов организмов в популяциях, смертность которых чрезвычайно высока на ранних стадиях развития, а в дальнейшем становится величиной постоянной, но достаточно высокой. Например, 1-й личиночный возраст у насекомых, при этом особи погибают в больших количествах, а гибель в последующих возрастных группах снижается.
· кривые промежуточного типа (II-III) относятся к тем видам, у которых смертность для каждой возрастной группы - величина постоянная.
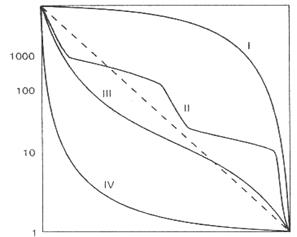 |
| По оси абцисс (горизонт.) - Возраст, в %% от продолжительности жизни По оси ординат (верт.) – число выживших на 1000 особей (или % выживших от исходной популяции). |
4. Типы кривых роста популяции
Экспоненциальная модель
Каждый вид стремится размножаться бесконечно (если его не сдерживают факторы среды).
Если среда не оказывает ограничивающего влияния, то специфическая скорость роста популяции для данных микроклиматических условий постоянна и максимальна.
В таких благоприятных условиях скорость роста характерна для некоторой возрастной структуры популяции и служит единственным показателем наследственно обусловленной способности популяции к росту.
| N0 | Количество особей в начальный момент времени | |||
| ∆N | Изменение числа организмов | |||
| ∆N / ∆t | Скорость роста (средняя скорость изменения числа организмов за определенное время) | 25 / год | ||
| ∆N / (N0∆t) | Удельная скорость роста (в расчете на одну особь). При умножении на 100 получаем скорость роста в %. | 25 / (50*1)=0.5 (или 50% в год) | ||
| r= dN / (Ndt) | Многновенная теоретическая скорость роста | 0.5 | ||
| dN / dt =rN | В данном уравнении подчеркивается, что скорость роста численности пропорциональна самой численности, а (r) - коэффициент пропорциональ-ности = 0,5 (в нашем случае) | |||
| dN/N=rdt | Интегрирование | |||
 | ||||
| Nt=N0ert | N0 – численность в начальный момент времени Nt – численность в момент времени t e = 2,7182 (Основание натуральных логарифмов) | |||
| lnNt=lnN0+rt | При логарифмировании | |||
 | Из уравнения ln nt= t - ln(N0) можно вычислить скорость роста популяции в начальный момент времени или в любой момент вермени, если вместо t подставить (t1-t2), а вместо N и N0 – (N1 и N2). r = const | |||
- Показатель r - фактическая разность между специфической мгновенной скоростью рождений (b) и специфической мгновенной скоростью гибели организмов (d). Она может быть выражена в виде:
r = b - d.
Экспоненциальный закон роста численности популяций был впервые установлен Мальтусом. Экспоненциальный рост характерен:
1) в эксперименте;
2) при заселении новых местообитаний;
3) для интродуцированных видов в благоприятных условиях.
- Уравнение J-образной кривой то же, что и при определении скорости роста. Разница состоит в том, что величина N имеет предел. Это означает, что относительно неограниченный рост внезапно останавливается, когда популяция исчерпает свои pecypcы (пища, жизненное пространство) или вмешается какой-либо друге фактор. После того как верхний предел N будет достигнут, плотность некоторое время может оставаться на этом уровне либо резко падать. Это характерно для природных популяций насекомых, водорослей и др.
· При определении стационарного и стабильного распределения возрастов специфическую скорость роста называют показателем потенциального роста популяции (rmax).
· Часто этот показатель называют биотическим, или репроду ктивным, потенциалом (термин введен Р. Чепменом в 1928 г.).
· Разность между биотическим потенциалом и фактической скоростью роста считают мерой сопротивления среды, которая характеризует сумму всех лимитирующих факторов, препятствующих реализации биотического потенциала.
Общая скорость роста популяции в отсутствие лимитирующего влияния среды зависит от возрастного состава и вклада в репродукцию различных возрастных групп. Следовательно, вид может характеризоваться величинами r в зависимости от структуры популяции.
Предположим, что популяция растет экспоненциально в течение времени равного длительности одного поколения (T). Тогда согласно уравнению кривой роста популяции мы получим:
NT=N0erT
Отсюда следует, что NT/N0 =erT
Однако, NT/N0, не что иное как отношение численности особей в двух следующих друг за другом поколениях, то есть R0 (смотри выше).
Таким образом, R0= erT. Из данного уравнения мы можем получить величину (r), как специфическую скорость роста популяции:
r = ln R0/ T
T – длительность поколения, которая определяется как промежуток между последовательными сменами генераций (от яйца до яйца).
· Смысл этой функции состоит в том, что через некоторое время рост численности замедляется. Она имеет вид S-образной кривой. Впервые была предложена как модель роста населения бельгийским исследователем Фэрхюльстом в 1938 г.
· В основе данной модели лежит предположение о линейном снижении скорости роста при увеличении численности. При этом скорость роста становится равной 0 при достижении некоторой максимальной численности.
- Когда численность стабилизируется вдоль емкости среды k происходят колебания, определяющиеся биологическими свойствами организма (продолжительность жизни организма, которая в целом пропорциональна его размеру, хотя есть много исключений).
При логистической (S-образной) кривой роста популяции вначале увеличение численности идет очень медленно, затем быстрее, но потом под влиянием факторов сопротивления среды рост популяции постепенно замедляется.
Это замедление, обусловливаемо сопротивлением среды, становится все более выраженным и в конечном итоге достигает определенной величины. Затем начинае поддерживаться более или менее устойчивое равновесие. Такой тип роста может быть выражен уравнением Ферхюльста-Пирла:

где К - константа, обозначающая верхний предел увеличения численности популяции, называемая верхней асимптотой для S-образно кривой.
Это уравнение отличается от экспоненциального тем, что содержит выражения (К - N) / К.
Это выражение представляет показатель, характеризующий сопротивление среды, создаваемое вследствие роста популяции, которая по мере приближения к пределу уменьшает скорость потенциальной репродукции.
Это уравнение отражает закон:скорость увеличения популяции равна максимально возможной скорости роста популяции, умноженной на степень реализации максимальной скорости.
Характерная форма S-образной кривой обусловлена постепенным усилением действия неблагоприятных факторов по мере увеличения плотности популяции.
В простом, или идеальном, случае усиление действия неблагоприятных факторов в зависимости от плотности популяции является линейным и его можно записать следующим образом:

где
r - скорость роста популяции или показатель потенциального роста,
N - величина популяции,
К - максимально возможная величина популяции;
е - основание натуральных логарифмов,
а - константа интегрирования, определяющая положение кривой относительно начала координат,
t - время.
Величина К называется предельной допустимой нагрузкой на среду, или емкостью среды для данной популяции.
Следует отметить, что для описания изменений численности популяции существует много математических уравнений, решение которых можно представить графически в виде S-образных кривых. Это справедливо почти для любого уравнения, в котором увеличение отрицательных факторов находится в какой-либо зависимости от плотности популяции.
Выбор математической модели определяется задачами исследований и адекватностью модели для каждого определенного случая.
5. Кривые роста популяций. Представление о емкости местообитания.
В зависимости от характера роста численности популяций выделяют различные типы их роста. По форме кривых, построенных на арифметической шкале, можно выделить два основных типа роста, описываемых J-образной и S-образной кривыми. Эти два типа кривых могут модифицироваться различным образом (рис. 1).

Рис. 1. Кривые роста численности популяции (по Ю. Одуму, 1875)
А - J-образная, В - S-образная форма кривой роста численности популяции.
A0 - вначале наблюдается неограниченный рост численности популяции;
А 1 - вначале наблюдается неограниченный рост численности, затем он прекращается и при благоприятных условиях вновь возобновляется, достигая прежней величины;
А2 - наблюдается нелимитированный рост численности, затем он внезапно прекращается и дальше наблюдаются колебания на более низком уровне;
В0 - происходит рост численности популяции по S-образной кривой, достигая К-уровня;
В1 - вначале наблюдается медленный рост численности, затем скорость возрастает и достигает К-уровня;
В2 - при достижении К-уровня наблюдаются небольшие отклонена от него;
В3 - наблюдаются значительные отклонения от К-уровня.
- Для сравнения экспериментальных данных с теоретической кривой следует убедиться в том, что показатели, входящие в уравнение, характеризуют воздействия, регулирующие плотность популяции.
- Ситуации, когда сопротивление среды возрастает линейно при увеличении плотности, могут иметь место в популяциях с простым жизненным циклом.
- В более высокоорганизованных популяциях, со сложным биологическим циклом и длительными периодами индивидуального развития, изменения, вероятно, отсрочены во времени.

