 2015-10-22
2015-10-22 9259
9259Выделительная система служит для выведения продуктов обмена и обеспечения водно-солевого состава организма. Основными органами выделения у рыб являются парные туловищные почки с их выводными протоками — мочеточниками, через которые моча поступает в мочевой пузырь. В некоторой степени в экскреции принимают участие кожа, жабры и кишечник.
Почки представляют собой систему выделительных канальцев, открывающихся в общий выводной проток.
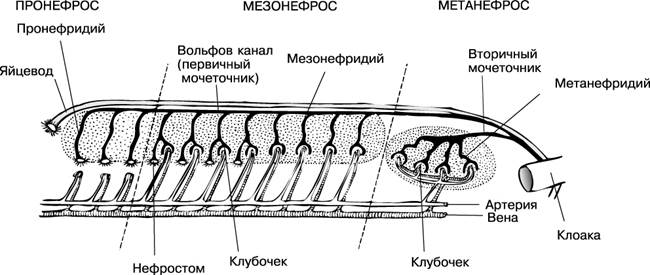
Эволюция выделительной системы у позвоночных происходит в последовательной смене трех типов почек: предпочки, туловищной и тазовой.
Предпочка, или головная почка (пронефрос), развивается в эмбриогенезе у низших позвоночных животных (круглоротые, рыбы, земноводные). Во взрослом состоянии у них функционирует первичная, или туловищная, почка (мезонефрос). У высших позвоночных (амниот) в эмбриогенезе функционирует туловищная почка, а у взрослых — вторичная, или тазовая (метанефрос). Прогрессивное развитие почек связано с усложнением строения почечных канальцев и редукцией мерцательных воронок.
Самой примитивной является головная почка, которая закладывается в передней части полости тела в виде 6-7 выделительных канальцев. Основным фильтрационным элементом ее является воронка, которая одним концом открывается в полость тела, а другим концом — в выводной проток — мочеточник. К верхней части воронки примыкает сосудистый клубочек. Такая почка функционирует у круглоротых и рыб на ранних стадиях развития. У некоторых рыб она сохраняется в виде 2-3 канальцев и во взрослом состоянии (бычки, атерина, бельдюга), а у большинства рыб преобразуется в лимфоидный орган, выполняющий функции кроветворения.
У взрослых круглоротых и рыб позади предпочки развиваются туловищные почки, лентовидные тяжи темно-красного цвета, заполняющие пространство между позвоночником и плавательным пузырем.
Основной функциональной единицей туловищной почки является нефрон, состоящий из мальпигиева тельца и выделительных канальцев. Мальпигиево тельце у рыб малоразмерное (диаметром 50-70 мкм), образовано капсулой Шумлянского — Боумена и сосудистым клубочком. Мочевые канальцы, отходящие от капсул, извиваются, а затем открываются в собирательные канальцы, которые объединяются в более крупные, впадающие затем в мочеточники (рис. 1).
У большинства рыб воронок в почках уже нет, они сохраняются лишь у некоторых хрящевых, а также у осетровых.
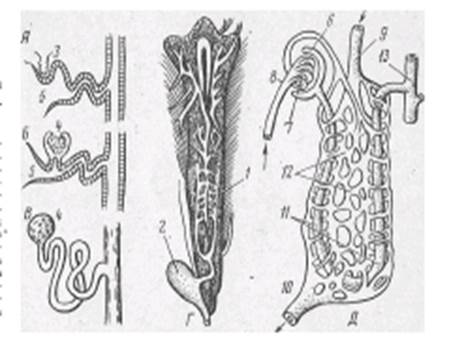 | Рис.1 Почка форели и схема почечных канальцев рыб: А — головная почка (пред- почка); Б, В, Д— туловищная почка; Г — почка форели; 1 — мочеточники; 2 — мочевой пузырь; 3 — наружный клубочек; 4 — мальпигиево тельце; 5 — воронка; 6 — капсула; 7 — сосудистый клубочек; 8 — ветвь спинной аорты; 9 — воротная вена почек; 10 — почечная вена; 11 — почечные канальцы; 12 — венозные синусоиды; 13 — собирательная трубка. |
 В почках выделяют три отдела: передний, средний и задний, которые у разных рыб имеют разную форму. У всех рыб передний отдел — головная почка. У сазана (карпа) в этом отделе правая и левая почки лежат отдельно, а в заднем срастаются в непарную ленту. Наиболее развит у них средний отдел, сильно расширенный и в виде больших лопастей охватывающий плавательный пузырь. У щуки и окуня передние и задние части почек слиты, а средние лежат раздельно.
В почках выделяют три отдела: передний, средний и задний, которые у разных рыб имеют разную форму. У всех рыб передний отдел — головная почка. У сазана (карпа) в этом отделе правая и левая почки лежат отдельно, а в заднем срастаются в непарную ленту. Наиболее развит у них средний отдел, сильно расширенный и в виде больших лопастей охватывающий плавательный пузырь. У щуки и окуня передние и задние части почек слиты, а средние лежат раздельно.
Основным компонентом мочи хрящевых рыб является мочевина, костистых — аммиак, причем аммиак токсичнее мочевины. Почки выполняют в основном роль фильтратора. Продукты обмена веществ доставляются в почки с кровью. От спинной аорты артериальная кровь по почечным артериям поступает в сосудистые клубочки, где происходит ее фильтрация и образуется первичная моча. Выходящие из сосудистых клубочков кровеносные сосуды вместе с сосудами воротной системы почек оплетают выделительные канальцы и, собираясь вместе, образуют задние кардинальные вены. В средней части канальцев происходит обратное всасывание веществ, нужных организму (воды, глюкозы, аминокислот), и образуется вторичная, или окончательная, моча.
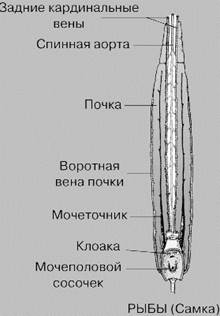
Выводным протоком головной почки является пронефрический канал. При развитии туловищной почки пронефрический канал расщепляется на два канала: вольфов и мюллеров. Мюллеров канал у самок хрящевых рыб выполняет функцию яйцевода, у самцов атрофируется. Вольфов канал у круглоротых, костистых и самок хрящевых рыб выполняет функцию мочеточника.

Рис. 2. Мочеполовая система круглоротых и некоторых рыб: I — миноги; II —акулы; III — осетровых; IV — лососевых; V — костистых; 1 — яичник; 2 — яйцевод (мюллеров канал); 3 — вторичный яйцевод; 4 — семенник; 5 — семяпровод; 6 — почка;
7 — мочеточник; 8 — клоака; 9 — мочеполовой синус.
У самцов хрящевых рыб на ранних стадиях развития вольфов канал выполняет одновременно функции мочеточника и семяпровода. У взрослых хрящевых образуется самостоятельный мочеточник, открывающийся в мочеполовой синус, а он в свою очередь — в клоаку, в то время как вольфов канал становится семяпроводом (рис. 2). Для мочеполовой системы костистых в отличие от других рыб характерно отсутствие у них клоаки и полное разделение выделительной и половой систем. Мочеточники (вольфовы каналы) у них по выходе из почек объединяются в непарный проток, который тянется вдоль задней стенки брюшной полости, образуя вырост — мочевой пузырь, заканчивающийся мочевым отверстием, которое открывается позади анального.

Рис. 3. Типы водно-солевого обмена костистых рыб (по Флорикену):
A— пресноводные; Б — морские костистые рыбы.
Строение почек и их функционирование у разных групп рыб связаны с особенностями осморегуляции. У морских хрящевых рыб кровь и тканевые жидкости изотоничны по отношению к окружающей среде, у пресноводных костистых гипертоничны, а у морских костистых гипотоничны. В связи с этим и осморегуляция у них осуществляется по-разному.
У морских хрящевых рыб изотоничность внутренней и внешней среды обеспечивается за счет удержания в крови и тканевых жидкостях мочевины и солей, в результате чего концентрация мочевины в крови у них достигает 2,0—2,5%. Клубочковый аппарат почек развит хорошо, но наружу выводятся лишь излишки мочевины, солей и воды, поэтому количество выделяемой мочи невелико (2—50 мл на 1 кг массы тела в сутки). Для выведения избытка солей у этих рыб есть особая ректальная железа, открывающаяся в прямую кишку.
В связи с тем что у пресноводных рыб осмотическое давление крови и тканевых жидкостей выше, чем в окружающей среде, вода проникает в организм через кожу, жабры, с пищей. Для предупреждения обводнения у них хорошо развит фильтрационный аппарат почек и выделяется большое количество мочи (50—300 мл на 1 кг массы тела в сутки). Потеря солей с мочой компенсируется активной реабсорбцией их в почечных канальцах и поглощением жабрами из воды (рис. 3).
Морские костистые рыбы живут в гипертонической среде, и вода выходит из организма через кожу, жабры, с мочой и фекалиями. Поэтому во избежание иссушения морские костистые рыбы пьют соленую воду, которая из кишечника всасывается в кровь. Часть солей из кишечника удаляется с фекалиями, другая часть выводится секреторными (хлоридными) клетками жаберного аппарата, и таким образом в жидкостях тела поддерживается относительно небольшое осмотическое давление. Клубочковая фильтрация развита слабо, и почки выводят небольшое количество мочи (0,5—20 мл на 1 кг массы тела в сутки). У некоторых рыб в процессе эволюции клубочки исчезают совсем (морская игла, морской черт).
Проходные рыбы при переходе из одной среды в другую могут изменять способ осморегуляции: в морской среде она осуществляется как у морских рыб, а в пресной — как у пресноводных, поддерживая осмотическое давление крови и тканевых жидкостей на определенном уровне.

