 2017-12-14
2017-12-14 2330
2330Рис. 1. Молекулярно-генетический уровень
1. Молекулярно-генетический уровень. Это наиболее элементарный характерный для жизни уровень (рис. 1). Как бы сложно или просто ни было строение любого живого организма, они все состоят из одинаковых молекулярных соединений. Примером этого являются нуклеиновые кислоты, белки, углеводы и другие сложные молекулярные комплексы органических и неорганических веществ. Их называют иногда биологическими макро- молекулярными веществами. На молекулярном уровне происходят различные процессы жизнедеятельности живых организмов: обмен веществ, превращение энергии. С помощью молекулярного уровня осуществляется передача наследственной информации, образуются отдельные органоиды и происходят другие процессы.
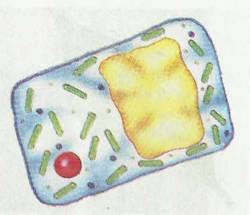
Рис. 2. Клеточный уровень
2. Клеточныйуровенъ. Клетка является структурной и функциональной единицей всех живых организмов на Земле (рис. 2). Отдельные органоиды в составе клетки имеют характерное строение и выполняют определенную функцию. Функции отдельных органоидов в клетке взаимосвязаны и выполняют единые процессы жизнедеятельности. У одноклеточных организмов (одноклеточные водоросли и простейшие) все жизненные процессы проходят в одной клетке, и одна клетка существует как отдельный организм. Вспомните одноклеточные водоросли, хламидомонады, хлореллу и простейших животных — амебу, инфузорию и др. У многоклеточных организмов одна клетка не может существовать как отдельный организм, но она является элементарной структурной единицей организма.

Рис. 3. Тканевый уровень
3. Тканевый уровень. Совокупность сходных по происхождению, строению и функциям клеток и межклеточных веществ образует ткань. Тканевый уровень характерен только для многоклеточных организмов. Также отдельные ткани не являются самостоятельным целостным организмом (рис. 3). Например, тела животных и человека состоят из четырех различных тканей (эпителиальная, соединительная, мышечная, нервная). Растительные ткани называются: образовательная, покровная, опорная, проводящая и выделительная. Вспомните строение и функции отдельных тканей.

Рис. 4. Органный уровень
4. Органный уровень. У многоклеточных организмов объединение нескольких одинаковых тканей, сходных по строению, происхождению и функциям, образует органный уровень (рис. 4). В составе каждого органа встречается несколько тканей, но среди них одна наиболее значительная. Отдельный орган не может существовать как целостный организм. Несколько органов, сходных по строению и функциям, объединяясь, составляют систему органов, например пищеварения, дыхания, кровообращения и т. д.

Рис. 5. Организменный уровень
5. Организменный уровень. Растения (хламидомонада, хлорелла) и животные (амеба, инфузория и т. д.), тела которых состоят из одной клетки, представляют собой самостоятельный организм (рис. 5). А отдельная особь многоклеточных организмов считается как отдельный организм. В каждом отдельном организме происходят все жизненные процессы, характерные для всех живых организмов, — питание, дыхание, обмен веществ, раздражимость, размножение и т. д. Каждый самостоятельный организм оставляет после себя потомство. У многоклеточных организмов клетки, ткани, органы и системы органов не являются отдельным организмом. Только целостная система органов, специализированно выполняющих различные функции, образует отдельный самостоятельный организм. Развитие организма, начиная с оплодотворения и до конца жизни, занимает определенный промежуток времени. Такое индивидуальное развитие каждого организма называется онтогенезом. Организм может существовать в тесной взаимосвязи с окружающей средой.

Рис. 6. Популяционно-видовой уровень
6. Популяционно-видовой уровень. Совокупность особей одного вида или группы, которая длительно существует в определенной части ареала относительно обособленно от других совокупностей того же вида, составляет популяцию. На популяционном уровне осуществляются простейшие эволюционные преобразования, что способствует постепенному появлению нового вида (рис. 6).

Рис. 7 Биогеоценотический уровень
7. Биогеоценотический уровень. Совокупность организмов разных видов и различной сложности организации, приспособленных к одинаковым условиям природной среды, называется биогеоценозом, или природным сообществом. В состав биогеоценоза входят многочисленные виды живых организмов и условия природной среды. В природных биогеоценозах накапливается энергия и передается от одного организма к другому. Биогеоценоз включает неорганические, органические соединения и живые организмы (рис. 7).

Рис. 8. Биосферный уровень
8. Биосферный уровень. Совокупность всех живых организмов на нашей планете и общей природной среды их обитания составляет биосферный уровень (рис. 8). На биосферном уровне современная биология решает глобальные проблемы, например определение интенсивности образования свободного кислорода растительным покровом Земли или изменения концентрации углекислого газа в атмосфере, связанные с деятельностью человека. Главную роль в биосферном уровне выполняют "живые вещества", т. е. совокупность живых организмов, населяющих Землю. Также в биосферном уровне имеют значение "биокосные вещества", образовавшиеся в результате жизнедеятельности живых организмов и "косных" веществ (т. е. условий окружающей среды). На биосферном уровне происходит круговорот веществ и энергии на Земле с участием всех живых организмов биосферы.
Уровни организации жизни. Популяция. Биогеоценоз. Биосфера.
1. В настоящее время выделяют несколько уровней организации живых организмов: молекулярный, клеточный, тканевый, органный, организменный, популяционно-видовой, биогеоценотический и биосферный.
2. На популяционно-видовом уровне осуществляются элементарные эволюционные преобразования.
3. Клетка — самая элементарная структурная и функциональная единица всех живых организмов.
4. Совокупность сходных по происхождению, строению и функциям клеток и межклеточных веществ образует ткань.
5. Совокупность всех живых организмов на планете и общей природной среды их обитания составляет биосферный уровень.
Вопрос 2
В растительной клетке выделяют протопласт (содержимое живой клетки) и его производные. В состав протопласта входят цитоплазма, ядро и другие органоиды: эндоплазматическая сеть, аппарат Гольджи, лизосомы, сферосомы, митохондрии, пластиды, рибосомы. К производным протопласта относят клеточную стенку, вакуоль и эргастические вещества.
Цитоплазма – многофазная высокоупорядоченная коллоидная система, заключенная между плазмолеммой и ядром. Цитоплазма представляет собой вязкую прозрачную бесцветную массу; упруга, эластична, с водой не смешивается. В молодой клетке ЦП занимает всю полость клетки, в старой – появляются вакуоли, сливающиеся со временем в одну крупную вакуоль, цитоплазма образует узкий постенный слой. В цитоплазме вода составляет 75-86%, белки – 10-20, липиды – 2-3, углеводы – 1-2, минеральные соли – 1%.
Ядро – важнейший компонент живой клетки. Впервые ядро было описано Р. Броуном в 1833 г. Ядро выполняет две важные функции: 1) контролирует жизнедеятельность клетки; 2) хранит генетическую информацию и передает ее дочерним клеткам в процессе деления. Форма ядра, как правило, округлая, но бывает веретеновидная, нитевидная, лопастная и др., размеры – от 2-3 до 500 мкм. Ядро состоит из ядерной оболочки, или мембраны, хроматиновых структур, ядрышка и ядерного сока.
Эндоплазматический ретикулум (ЭР) – органоид клетки, представляющий собой систему мелких вакуолей и канальцев, соединенных друг с другом и ограниченных одинарной мембраной. Мембраны ЭР толщиной 5-7 нм нередко переходят в ядерную мембрану. Различают два типа ЭР – шероховатый (гранулярный, несет на своих мембранах рибосомы) и гладкий (агранулярный, лишен рибосом). Гранулярный ЭР связывает в единое целое все структурно- функциональные единицы клетки, обеспечивает транспорт ионов и макромолекул внутри клетки, а также синтез белков на прикрепленных Ботаника. Клетка и ткани: практ. рук-во / Ю. М. Бачура, Н. М. Дайнеко рибосомах. Гладкий ЭР участвует в синтезе липидов, обмене некоторых полисахаридов, накоплении и выведении из клетки ядовитых веществ.
Митохондрии – округлые или цилиндрические, реже нитевидные двухмембранные органеллы длиной до 10 мкм, диаметром 0,2-1 мкм. Внутренняя мембрана образует выросты – кристы, которые в растительных клетках обычно имеют вид трубочек. Внутри митохондрии заполнены матриксом, в котором содержатся молекулы митохондриальной ДНК, РНК и рибосомы. Основная функция митохондрий – образование энергии.
Аппарат Гольджи (комплекс Гольджи) состоит из отдельных диктиосом и пузырьков Гольджи.
Диктиосомы – органеллы, представляющие собой пачки (2-7 и более) плоских округлых цистерн, ограниченных мембраной и заполненных матриксом. В цистернах аппарата Гольджи накапливаются, конденсируются и упаковываются вещества, подлежащие изоляции или удалению из цитоплазмы. Упакованные в пузырьки, они поступают в вакуоли.
Аппарат Гольджи – место синтеза полисахаридов, идущих на построение клеточной стенки. Пузырьки Гольджи участвуют также в формировании новых клеточных стенок и плазмалеммы, происходящем после митоза.
Рибосомы – мельчайшие клеточные органеллы, около 17-23 нм в диаметре, состоящие примерно из равных количеств белка и нуклеиновых кислот. Рибосомы располагаются в цитоплазме свободно или связаны с мембранами эндоплазматической сети. Рибосомы состоят из двух субъединиц: крупной округлой и мелкой несколько сплющенной. Роль рибосом – внутриклеточный синтез белка.
Микротельца – сферические или палочковидные мелкие (0,2-1,5 мкм) одномембранные органеллы с плотным матриксом, состоящим в основном из окислительно-восстановительных ферментов. Различают: 1) пероксисомы – микротельца, играющие важную роль в метаболизме гликолевой кислоты и имеющие непосредственное отношение к фотодыханию; 2) глиоксисомы – микротельца, содержащие ферменты необходимые для превращения жиров в углеводы.
Лизосомы – округлые одномембранные органеллы, в матриксе которых содержится большое число гидролитических ферментов. Лизосомы осуществляют внутриклеточное переваривание, автолиз. Гидролитические ферменты лизосом очищают всю полость клетки после отмирания ее протопласта (например, при образовании сосудов).
Пластиды – органеллы, характерные исключительно для растительных клеток. Форма, размеры, строение и функции пластид различны. Они окружены двойной мембраной и заполнены матриксом. В матриксе имеются кольцевая ДНК и рибосомы прокариотического Ботаника. Клетка и ткани: практ. рук-во / Ю. М. Бачура, Н. М. Дайнеко типа. Различают три типа пластид: хлоро-, хромо- и лейкопласты. Хлоропласты высших растений имеют примерно одинаковую форму двояковыпуклой линзы. Их размеры: 5-10 мкм в длину при диаметре 2-4 мкм. Число хлоропластов в клетках высших растений 15- 50. Внутренняя мембрана хлоропластов образует в строме систему замкнутых карманов – тилакоидов. Группы тилакоидов образуют стопки – граны. Хлоропласты часто содержат зерна крахмала, липиды. В онтогенезе хлоропласты формируются из пропластид путем образования из впячиваний внутренней мембраны уплощенных мешков – тилакоидов. Функция хлоропластов – фотосинтез; также они могут участвовать в синтезе аминокислот, служить хранилищем временных запасов крахмала.
Лейкопласты – бесцветные округлые пластиды, в которых обычно накапливаются запасные питательные вещества, в основном крахмал. По строению лейкопласты мало отличаются от пропластид, из которых они образуются: двумембранная оболочка окружает бесструктурную строму. Внутренняя мембрана, врастая в строму, образует немногочисленные тилакоиды. Лейкопласты, в которых синтезируется и накапливается запасной крахмал, называются амилопластами, белки – протеинопластами, масла – элайопластами.
Хромопласты – пластиды оранжево-красного и желтого цвета, образующиеся из лейкопластов и хлоропластов в результате накопления в их строме каротиноидов. Они встречаются в клетках лепестков (лютик, нарцисс, тюльпан, одуванчик), зрелых плодов (томат, тыква, арбуз, апельсин), редко – корнеплодов (морковь, кормовая свекла), а также в осенних листьях. Хромопласты – конечный этап в развитии пластид. Косвенное биологическое значение хромопластов в том, что ярко окрашенные плоды успешнее распространяются птицами и животными, а выделяющиеся яркой желто-красной окраской цветки привлекают насекомых-опылителей.
Вопрос 3
Геном современных прокариотических клеток характеризуется относительно небольшими размерами. У кишечной палочки (Е. coli) он представлен кольцевой молекулой ДНК длиной около 1 мм, которая содержит 4·106 пар нуклеотидов, образующих около 4000 генов. Основная масса ДНК прокариот (около 95%) активно транскрибируется в каждый данный момент времени. Как было сказано выше, геном прокариотической клетки организован в виде нуклеоида — комплекса ДНК с негистоновыми белк а ми
В прокариотических (бактериальных) клетках обнаружены плазмиды, которые несут наследственный материал, определяющий такие свойства, как способность бактерий к конъюгации, а также их устойчивость к некоторым лекарственным веществам.
Вопрос 4
Хромосомы - это нуклеопротеидные структуры, располагающиеся в ядрах эукариотических клеток. В них хранится практически вся наследственная информация, и именно они несут функцию ее хранения, передачи и реализации. Хромосомы практически не видны даже в световой микроскоп, однако их можно четко рассмотреть в периоды деления клетки, во время митоза и мейоза. Кариотип и правила хромосом Кариотипом называется совокупность всех хромосом (диплоидный набор), находящихся в клетке. Он видоспецифичен, то есть является уникальным для каждого вида живых существ на планете, уровень изменчивости его относительно низок, однако у некоторых особей может обладать определенными особенностями. Например, представители разного пола имеют в основном одинаковые хромосомы (аутосомы), различие кариотипов составляет только одна пара хромосом - половые хромосомы, или гетерохромосомы. Правила хромосом просты: число их постоянно (в соматических клетках может содержаться только строгое число хромосом, например, у кошек - 38, у плодовой мушки дрозофилы - 8, у курицы - 78, а у человека 46). Хромосомы парны, каждая из них имеет гомологичную пару, идентичную по всем параметрам, включая форму и размер. Разнится только происхождение: одна - от отца, другая - от матери. Гомологичные пары хромосом индивидуальны: каждая из пар отличается от других не только внешним видом - формой и размером, - но и расположением светлых и темных полос. Непрерывность - еще одно правило хромосом. ДНК клетки удваивается перед делением, результатом чего становится пара сестринских хроматид. Каждая дочерняя клетка после деления получает по одной хроматиде, то есть от хромосомы образуется хромосома. Необходимые элементы Хромосома, строение которой относительно несложно, образуется из молекулы ДНК, обладающей большой длиной. Она содержит линейные группы множества генов. Каждая хромосома обладает центромерой и теломерами, точками инициации репликации - это ее необходимые функциональные элементы. Теломеры находятся на кончиках хромосом. За счет них и точек начала репликации (их так же называют сайтами инициации), молекула ДНК может реплицироваться. В центромерах же происходит прикрепление сестринских молекул ДНК к митотическому веретену деления, что позволяет им точно разойтись по дочерним клеткам во время процесса митоза.
Химический состав хромосом
Кратко можно описать, что наследственный материал в хромосомах предстает как нуклео-протеиновый комплекс. После изучения химической организации хромосом в эукариотических клетках, ученые могут сказать, что состоят они в большей части из ДНК и белков, которыми образуется нуклео-протеиновый комплекс, называемый хроматином. Белки, входящие состав хромосом, это значительная часть всего вещества в хромосомах, около 65% всей массы структур приходится именно на них. Хромосомные белки подразделяются на негистоновые белки и гистоны. Гистоны - сильноосновны, щелочной характер их обуславливается наличием лизина и аргенина - основных аминокислот. Химический и структурный состав хромосом разнообразен. Гистоны представляют пять фракций: Hl, H2A, H2B, H3 и H4. Все, кроме первой фракции, примерно в равных количествах имеются в клетках всех видов, принадлежащих к высшим млекопитающим. Белков Hl меньше вдвое. Синтез гистонов происходит на полисомах цитоплазмы. Это основные белки, имеющие положительный заряд, за счет чего могут прочно соединяться с молекулами ДНК и таким образом не дают считывать заключенную наследственную информацию. В этом заключается регуляторная роль гистонов, но помимо нее есть и структурная функция, за счет которой обеспечивается пространственная организация ДНК в хромосомах. В характерный химический состав интерфазных хромосом входят и негистоновые белки, которые, в свою очередь, подразделяются более чем на сто фракций. В этот ряд входят ферменты, отвечающие за синтез РНК, и ферменты, которые запускают репарацию и редупликацию ДНК. Так же как и основные, кислые хромосомные белки имеют регуляторную и структурную функции. Однако химический состав хромосомы на этом не заканчивается: кроме белков и ДНК, в составе присутствует РНК, ионы металлов, липиды и полисахариды. Отчасти хромосомная РНК присутствует в качестве продуктов транскрипции, которые еще не покинули место синтеза.
По положению центромер и длине плеч, располагающихся по обе стороны от нее, хромосомы классифицируются на метацентрические (равноплечие), если центромера располагается посредине, и субметацентрические (неравноплечие), если центромера сдвинута к одному из концов. Также существуют акроцентрические, или палочковидные хромосомы (центромера у них расположена практически на самом конце) и точковые хромосомы, получившие свое название за небольшой размер, вследствие чего практически невозможно определить их форму. У телоцентрических хромосом тоже трудно определить место расположения первичной перетяжки.
Свойства и функции генов
Ген считается элементарной неделимой функциональной единицей наследственного матриала. Она имеет вид участка молекулы ДНК, которой кодируется структура как минимум одного пептида. Ген имеет определенные свойства, первое из них - дискретность действия. Это означает, что различно локализованные гены контролируют развитие признаков особи. Свойство постоянства определяется тем, что ген неизменен при наследственной передаче, если, конечно, не произошло мутации. Из этого следует, что ген не может быть изменен в течение жизни. Специфичность действия заключается в обусловленности развития признака или группы признаков, однако гены могут оказывать и множественные действия - это называется плейотропией. Свойство дозированности действия определяет предел, до которого может развиться признак, обусловленный геном. Для них также характерно и аллельное состояние, то есть практически все гены находятся в аллелях, количество которых начинается с двух.
Вопрос 5
Исходным моментом для понимания явлений наследственности служат данные о химическом составе хромосом, представляющих собой индивидуальные компоненты клеточных ядер. Главным в химической характеристике хромосом служит то, что они являются комплексными полимерами из белков и нуклеиновых кислот.
Доказано, что генетическая информация организмов, созданная в течение их эволюции, кодирована в молекулярных структурах ДНК, входящих в состав хромосом клетки, или в структурах нитей ДНК (в ряде случаев РНК), свойственных бактериям и вирусам.
Молекулы ДНК, по теории Уотсона и Крика (1953), состоят из двух цепей полинуклеотидов. Каждый нуклеотид содержит пентозу, соединенную с фосфатом и с одним азотистым основанием. Всего гетероциклических оснований в ДНК четыре: аденин, тимин, гуанин и цитозин. Цепь повторяющихся пентоз и фосфатов составляет основу полинуклеотидной цепи в молекуле ДНК. Разнообразие участков молекулы возникает за счет своеобразия взаимоположения разных нуклеотидов. Две цепи ДНК соединены в молекулу благодаря водородным связям, соединяющим попарно (из разных цепей) аденин (А) — тимин (Т), гуанин (Г) — цитозин (Ц). Эта система обеспечивает и механизм авторепродукции молекул. При разрыве водородных связей спираль молекулы ДНК развертывается. Освобождающиеся цепи служат матрицей для синтеза дочерней спирали, которая становится комплементарной (А притягивается только к Т и, наоборот, то же и для Г - Ц). В результате обе вновь образовавшиеся двойные спирали молекул ДНК оказываются идентичными исходной спирали.
В этом механизме авторепродукции заключены основы консерватизма наследственности. Корнберг в экспериментальных условиях обнаружил, что при наличии нуклеозидтрифосфатов и особого фермента — полимеразы внесение в раствор небольшого количества ДНК в качестве затравки приводит к тому, что синтезируемая в этих условиях ДНК строго воспроизводит именно то соотношение и взаиморасположение аденина, тимина, цитозина и гуанина, которое было свойственно молекуле «затравки». В механизме авторепродукции молекул ДНК заложен также механизм воспроизведения мутаций в неограниченном ряду поколений.
Само явление генных мутаций связано с изменениями в составе или в порядке оснований в определенном локусе (лат. locus — место) молекулы ДНК. Вполне понятно, что, раз возникнув, такое изменение в химической структуре ДНК будет затем по законам авторепродукции передаваться всем последующим синтезируемым молекулам.
Молекулярные основы наследственности, т. е. кодирование генетической информации в химической структуре нуклеиновых кислот, принципиально одинаковы у всех форм жизни на Земле. Однако преемственность организмов связана с воспроизведением наследственных структур клетки, в которых молекулы ДНК оказываются включенными в надмолекулярную мицеллярную микроскопическую организацию в виде хромосом. Исключением являются вирусы, однако и для них процесс воспроизведения неизвестен вне клеток хозяина. В этом случае для синтеза новых молекул ДНК или РНК вирусов используется система клетки хозяина.
Исследования Тейлора (1963) показали, что авторепродукция хромосом совершается по полуконсервативному типу. В первой метафазе после введения тимидина, меченного радиоактивным тритием, обе хроматиды несут метку. Во второй метафазе одна хроматида оказывается свободной от метки, а другая несет метку. Это показывает, что авторепродукция хромосомы совершается по тем же принципам, что и авторепродукция молекулы ДНК по схеме Уотсона и Крика. Исходя из этого, Тейлор предлагает схему строения основной нерасщепленной хромосомы из одной непрерывной нити ДНК, сложно упакованной в белковой оболочке.
Сложная организация хромосомы, и в первую очередь обязательное наличие в ней белка, имеет принципиальное значение. Белок обладает защитными функциями и может иметь большое значение при репарациях в пораженных молекулах ДНК. Спирализация хромосом связана с количеством гистонов. Форма организации хромосом у вирусов и бактерий, с одной стороны, и у всех остальных форм — с другой, заметно различаются. У первых наследственные структуры представлены непосредственно молекулами ДНК, у остальных форм молекулы ДНК связаны хромосомными белками.
Каждая хромосома, входящая в ядро клеток любого вида, обладает качественной генетической индивидуальностью, составляя важнейшую часть в общей клеточной организации.
Генетический материал хромосом (ДНК) регулирует жизнедеятельность клетки и сам строится в процессах обмена веществ на базе законов авторепродукции, установленных для молекул ДНК.
Вопрос 6
3. Деление клетки
Способность к делению — это важнейшее свойство клетки. В результате деления из одной клетки возникают две новые. Одно из основных свойств жизни — самовоспроизведение — проявляется уже на клеточном уровне. Наиболее распространенным способом деления клетки является митоз — непрямое деление клетки.
Митоз — это процесс образования двух дочерних клеток с набором хромосом, идентичным исходной материнской клетке. Митотическое деление приводит к увеличению числа клеток, обеспечивает рост организма, регенерацию или замену клеток в процессе их старения. У некоторых организмов митоз лежит в основе их размножения бесполым путем.
Деление клетки состоит из двух последовательных процессов: кариокинеза — деления ядра, или собственно митоза, и цитокинеза — деления цитоплазмы.
В процессе кариокинеза происходит основное, наиболее важное событие — перераспределение хромосом, т. е. молекул ДНК, обеспечивающее равномерную передачу наследственной информации между двумя дочерними клетками.
В процессе цитокинеза осуществляется более-менее равномерное распределение цитоплазмы и ее органоидов между двумя дочерними клетками. Однако это событие не происходит с такой точностью, как процесс кариокинеза. События, происходящие в митозе, можно увидеть в световой микроскоп на фиксированных препаратах. Современные методы фазовоконтрастной микроскопии и микрофотосъемки дали возможность наблюдать этот процесс в живой клетке.
В настоящее время клеточный цикл и митоз изучаются на отдельных изолированных клетках. Клеточную популяцию, полученную от одной исходной материнской клетки, называют клоном.
Рассмотрим подробно процессы, происходящие в период деления.
Фазы митоза
Митоз состоит из четырех последовательных фаз, обеспечивающих равномерное распределение генетической информации и всех органоидов между двумя дочерними клетками (рис. 5).
Профаза — первая, самая продолжительная фаза митоза. Переход из фазы G интерфазы в профазу митоза происходит постепенно. Хроматин начинает уплотняться, и формируются хромосомы. Они максимально спирализуются, утолщаются и становятся хорошо заметными. Каждая хромосома состоит из двух сестринских хроматид, состоящих из молекулы ДНК. В этот период жизнедеятельности клетки количество хромосом в диплоидной клетке составляет 2n, а число молекул ДНК удвоено и равно 4с. Сестринские хроматиды соединены друг с другом центромерой. Ядерная мембрана рассасывается, ядрышко распадается, все органоиды перестают различаться в световой микроскоп.
Центриоли клеточного центра расходятся к полюсам, образуя веретено деления, состоящее из микротрубочек. Микротрубочки располагаются вокруг центриолей в виде звезды. В конце профазы ядерная мембрана исчезает полностью.
Рис. 5. Стадии митоза: 1 — профаза; 2 — метафаза; 3 — анафаза; 4 — телофаза
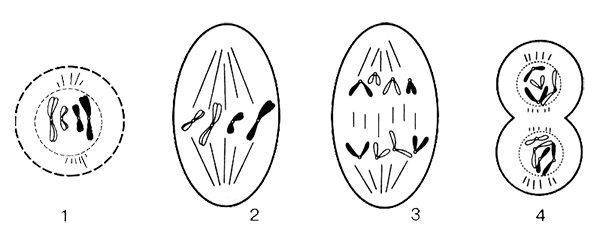
Рис. 6. Схема митотического веретена деления в метафазе: 1 — хромосома; 2 — микротрубочки веретена делении; 3 — зона перекрывания микротрубочек; 4 — центриоли

Метафаза. Во вторую стадию митоза нити веретена деления соединяются с центромерами хромосом и перемешают их в экваториальную зону клетки. В конце метафазы все хромосомы выстраиваются в одной экваториальной плоскости. В этом положении они удерживаются микротрубочками веретена деления. Микротрубочки связаны с центромерами хромосом. Они обеспечивают движение хромосом к экватору и их выстраивание на равном расстоянии от полюсов клетки (рис. 6).
Анафаза — достаточно короткая стадия. Сестринские хроматиды разделяются в местах центромеры. Каждая центромера делится, и хромосома распадается на две сестринские хроматиды, которые теперь становятся отдельными хромосомами. Начинается движение сестринских хроматид — хромосом к полюсам клетки, которые растягиваются нитями веретена деления. У каждого полюса оказывается такое же количество хромосом, которое было в исходной материнской клетке.
Телофаза. В последнюю фазу кариокинеза происходит формирование новых ядер у полюсов клетки. Хромосомы деспирализуются, нити веретена деления исчезают. Вновь формируется ядрышко. Эта фаза завершается цитокинезом.
Цитокинез
Процесс деления ядра плавно переходит в процесс деления цитоплазмы. Клеточные органоиды равномерно распределяются по двум полюсам клетки. В экваториальной части плазматическая мембрана образует впячивание, которое втягивается внутрь клетки. Полагают, что этот процесс связан с деятельностью микротрубочек. В клетке образуется борозда деления — перетяжка из клеточной мембраны, которая постепенно углубляется к центру клетки. Возникают две новые дочерние клетки, идентичные исходной материнской.
Весь процесс деления длится от нескольких минут до трех часов, в зависимости от типа клеток и организма. Митоз в несколько раз по времени короче интерфазы.
В растительных и животных клетках митоз имеет ряд отличий. Во-первых, в растительных клетках центриоли отсутствуют. Во-вторых, нити веретена деления исчезают не полностью, а сохраняются в экваториальной зоне, где за счет содержимого пузырьков аппарата Гольджи образуется клеточная пластинка. Разрастаясь, она сливается со стенками исходной клетки и разделяет материнскую клетку на две дочерние. Мембраны пузырьков идут на построение новых клеточных мембран. Таким образом, впячивания и перетяжки здесь не образуется. За счет целлюлозных волокон, которые придают клетке прочность и эластичность, происходит формирование первичной клеточной стенки. Постепенно в процессе роста клеточная стенка изменяется, утолщается за счет отложения целлюлозных волокон. Она приобретает слоистую структуру, в ней образуются поры, пронизанные плазмодесмами.
Биологический смысл митоза заключается в обеспечении постоянства числа хромосом и идентичности наследственной информации вновь возникающих клеток из исходной материнской клетки. Митоз обеспечивает генетическую стабильность клетки.
Амитоз
Иногда встречается и другой вид деления клетки — амитоз. Амитоз — это прямое деление ядра, без образования хромосом и веретена деления. При этом наследственная информация распределяется неравномерно. Амитоз встречается у некоторых простейших, в клетках специализированных тканей (хрящи), в раковых клетках.
Вопрос 7
Митоз, кариокинез, или непрямое деление, — универсальный, широко распространенный способ деления клеток. При этом конденсированные и уже редуплицированные хромосомы переходят в компактную форму митотических хромосом, образуется веретено деления, участвующее в сегрегации и переносе хромосом (ахроматиновый митотический аппарат), происходит расхождение хромосом к противоположным полюсам клетки и деление тела клетки (цитокинез, цитотомия).Процесс непрямого деления клеток принято подразделять на несколько основных фаз: профаза, метафаза, анафаза, телофаза.
Митоз (кариокинез).
Клеточный цикл – это жизнь клетки от одного митотического деления до другого.
Деление ядра – кариокинез. Деление цитоплазмы – цитокинез.
Клеточный цикл состоит из 3х стадий:
1. Интерфаза. Период интенсивного синтеза веществ, роста и развития клетки.
Состоит из 3х периодов:
1. Период G1: активный синтез НК, белков, жиров, углеводов; формируются органеллы; синтезируются вещества, регулирующие следующую фазу цикла.
2. Период S: редупликация ДНК
3. Период G2: биосинтез специальных белков (для веретена деления), синтезируются АТФ.
2. Собственно митоз.
1. Профаза.
- Хромосомы уплотняются и становятся видимы;
- сестринские хромосомы соприкасаются;
- исчезают ядрышки;
- органоиды расходятся к периферии клетки;
- образуется веретено деления;
2. Метафаза.
1. Хромосомы выстраиваются на экваторе
3. Анафаза.
2. Расщепляются центромеры;
3. Сестринские хромосомы расходятся к полюсам.
4. Телофаза.
- Хромосомы деспирализуются → хроматин
- Образуется ядерная оболочка
- Формируются ядрышки
- Разрушаются нити веретена деления
Биологическая сущность:
- Обеспечение генетической стабильности
- Увеличение числа клеток → рост и развитие тканей
- Бесполое размножение
- Регенерация тканей
Вопрос 8
При половом размножении дочерний организм возникает в результате слияния двух половых клеток (гамет) и последующего развития из оплодотворенной яйцеклетки — зиготы.
Половые клетки родителей обладают гаплоидным набором (n) хромосом, а в зиготе при объединении двух таких наборов число хромосом становится диплоидным (2 n): каждая пара гомологичных хромосом содержит одну отцовскую и одну материнскую хромосому.
Гаплоидные клетки образуются из диплоидных в результате особого клеточного деления — мейоза.
Мейоз — разновидность митоза, в результате которого из диплоидных (2п) соматических клеток половых желез образуются гаплоидные гаметы (1n). При оплодотворении ядра гаметы сливаются, и восстанавливается диплоидный набор хромосом. Таким образом, мейоз обеспечивает сохранение постоянного для каждого вида набора хромосом и количества ДНК.
Мейоз представляет собой непрерывный процесс, состоящий из двух последовательных делений, называемых мейозом I и мейозом II. В каждом делении различают профазу, метафазу, анафазу и телофазу. В результате мейоза I число хромосом уменьшается вдвое (редукционное деление): при мейозе II гаплоидность клеток сохраняется (эквационное деление). Клетки, вступающие в мейоз, содержат генетическую информацию 2n2хр (рис. 1).
В профазе мейоза I происходит постепенная спирализация хроматина с образованием хромосом. Гомологичные хромосомы сближаются, образуя общую структуру, состоящую из двух хромосом (бивалент) и четырех хроматид (тетрада). Соприкосновение двух гомологичных хромосом по всей длине называется конъюгацией. Затем между гомологичными хромосомами появляются силы отталкивания, и хромосомы сначала разделяются в области центромер, оставаясь соединенными в области плеч, и образуют перекресты (хиазмы). Расхождение хроматид постепенно увеличивается, и перекресты смещаются к их концам. В процессе конъюгации между некоторыми хроматидами гомологичных хромосом может происходить обмен участками — кроссинговер, приводящий к перекомбинации генетического материала. К концу профазы растворяются ядерная оболочка и ядрышки, формируется ахроматиновое веретено деления. Содержание генетического материала остается прежним (2n2хр).
В метафазе мейоза I биваленты хромосом располагаются в экваториальной плоскости клетки. В этот момент спирализация их достигает максимума. Содержание генетического материала не изменяется (2п2хр).
В анафазе мейоза I гомологичные хромосомы, состоящие из двух хроматид, окончательно отходят друг от друга и расходятся к полюсам клетки. Следовательно, из каждой пары гомологичных хромосом в дочернюю клетку попадает только одна — число хромосом уменьшается вдвое (происходит редукция). Содержание генетического материала становится 1n2хр у каждого полюса.
В телофазе происходит формирование ядер и разделение цитоплазмы — образуются две дочерние клетки. Дочерние клетки содержат гаплоидный набор хромосом, каждая хромосома — две хроматиды (1n2хр).
Интеркинез — короткий промежуток между первым и вторым мейотическими делениями. В это время не происходит репликации ДНК, и две дочерние клетки быстро вступают в мейоз II, протекающий по типу митоза.

Рис. 1. Схема мейоза (показана одна пара гомологичных хромосом). Мейоз I: 1, 2, 3. 4. 5 — профаза; 6 —метафаза; 7 — анафаза; 8 — телофаза; 9 — интеркинез. Мейоз II; 10 —метафаза; II —анафаза; 12 — дочерние клетки.
В профазе мейоза II происходят тс же процессы, что и в профазе митоза. В метафазе хромосомы располагаются в экваториальной плоскости. Изменений содержания генетического материала не происходит (1n2хр). В анафазе мейоза II хроматиды каждой хромосомы отходят к противоположным полюсам клетки, и содержание генетического метериала у каждого полюса становится lnlxp. В телофазе образуются 4 гаплоидные клетки (lnlxp).
Таким образом, в результате мейоза из одной диплоидной материнской клетки образуются 4 клетки с гаплоидным набором хромосом. Кроме того, в профазе мейоза I происходит перекомбинация генетического материала (кроссинговер), а в анафазе I и II — случайное отхождение хромосом и хроматид к одному или другому полюсу. Эти процессы являются причиной комбинативной изменчивости.
Биологическое значение мейоза:
1) является основным этапом гаметогенеза;
2) обеспечивает передачу генетической информации от организма к организму при половом размножении;
3) дочерние клетки генетически не идентичны материнской и между собой.
Атак же, биологическое значение мейоза заключается в том, что уменьшение числа хромосом необходимо при образовании половых клеток, поскольку при оплодотворении ядра гамет сливаются. Если бы указанной редукции не происходило, то в зиготе (следовательно, и во всех клетках дочернего организма) хромосом становилось бы вдвое больше. Однако это противоречит правилу постоянства числа хромосом. Благодаря мейозу половые клетки гаплоидны, а при оплодотворении в зиготе восстанавливается диплоидный набор хромосом (рис. 2 и 3).

Рис. 2. Схема гаметогенеза:? — сперматогенез;? — овогенез
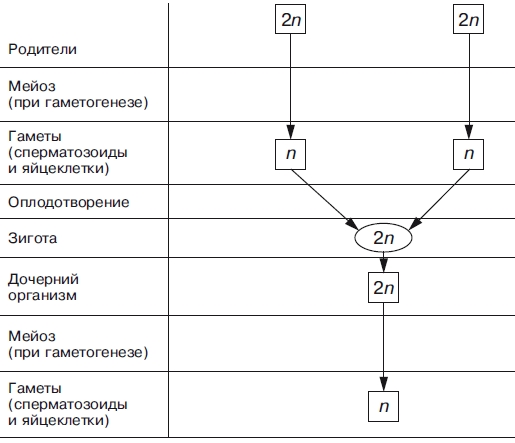
Рис. 3. Схема, иллюстрирующая механизм сохранения диплоидного набора хромосом при половом размножении
Вопрос 9
Типы размножения организмов, их классификация
Размножение, или воспроизведение себе подобных, — специфическое и обязательное свойство всех живых существ. Время индивидуальной жизни отдельных организмов весьма ограничено. Но благодаря их самовоспроизведению обеспечивается длительное существование как конкретных видов, так и живого в целом. Размножение, компенсируя процесс естественного отмирания особей, сохраняет вид в бесчисленном ряду поколений.
Самовоспроизведение основано на передаче от родителей к потомкам наследственной информации о совокупности признаков, свойств и качеств, присущих представителям данного вида. В ходе развития происходила эволюция и форм размножения, разнообразием которых отличаются современные живые организмы.
Классификация способов размножения. Прежде всего необходимо выделить два принципиально отличающихся типа размножения организмов — бесполое и половое (рис. 1).

Рис. 1. Сравнительные схемы двух основных типов размножения:? — бесполое размножение (одна особь производит двух или
большее число потомков);? — половое размножение (две гаметы от двух родительских особей, соединяясь, дают начало новому организму)
При бесполом размножении из одной клетки (или группы клеток у многоклеточных) родительского организма при ее миотическом делении образуется новая особь. Поэтому образующиеся дочерние организмы сходны друг с другом и со своим родителем по всем признакам. Образно говоря, в процессе бесполого размножения осуществляется «тиражирование» множества генетических копий родительского организма.
В половом размножении участвуют две родительские особи. Они образуют специализированные половые клетки — гаметы, в результате слияния которых (оплодотворения) возникает зигота (Z), дающая начало дочернему организму.
При образовании зиготы происходит комбинирование наследственной информации (объединение хромосомных наборов родителей). Следовательно, развивающийся из зиготы дочерний организм обладает новым сочетанием признаков. Таким образом, половое размножение обеспечивает разнообразие особей данного вида, что способствует освоению ими различных условий обитания, определяет комбинативную изменчивость. Именно этим объясняется преимущественное распространение полового процесса в разных царствах живых существ. Тем не менее у многих видов организмов при наличии полового процесса в ходе их жизненного цикла сохраняются и разные формы бесполого размножения. Это объясняется тем, что последнее может обеспечить быстрое и существенное увеличение численности особей при благоприятных условиях среды. Успех существования многих видов организмов обусловлен сочетанием разных способов их размножения (схема 1).
Схема 1. Способы размножения организмов
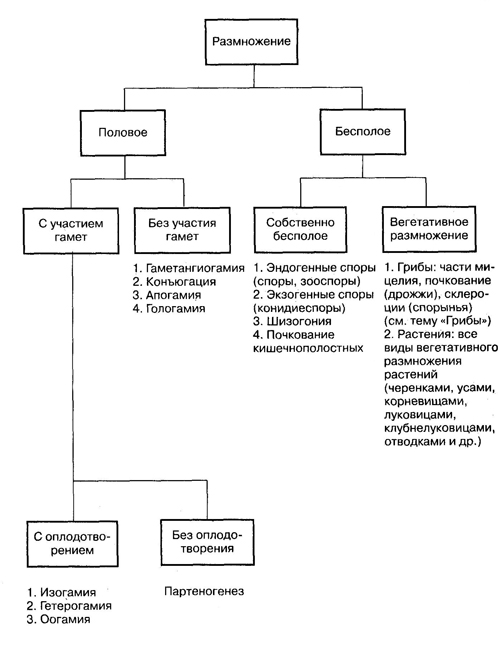
Источник: Краснодембский Е. Г."Общая биология: Пособие для старшеклассников и поступающих в вузы"
Вопрос 10 и 11
Процесс формирования половых клеток у растений подразделяется на два этапа: 1-й этап — спорогенез — завершается образованием гаплоидных клеток — спор, в ходе 2-го этапа — гаметогенеза — происходит ряд делений гаплоидных клеток, прежде чем образуются зрелые гаметы.
Процесс образования микроспор, или пыльцевых зерен, у растений называют микроспорогенезом, а процесс образования мегаспор (или макроспор) — мега- или макроспорогенезом. Микроспорогенез протекает аналогично делению созревания у животных мужских половых клеток до стадии сперматиды, а мегаспорогенез — соответственно до стадии незрелой яйцеклетки — ооцита II.
Процесс гаметогенеза у растений в принципе сходен с таковым у животных, но протекает несколько отличным путем. У животных после двух мейотических делений формируются гаметы, и никаких дополнительных клеточных делений не происходит. У растений в результате двух мейотических делений возникает гаплоидная спора, из которой развивается гаметофит, представляющий собой у низших растений (грибов, печеночников, мхов, ряда водорослей) целый организм и наиболее продолжительную стадию цикла существования. У высших растений гаплоидная фаза редуцирована, однако ядра мужской и женской спор претерпевают ряд митотических делений, прежде чем образуются гаметы.

