2014-01-31
2014-01-31 803
803Рис. 12
Рис. 11
Закон толерантности – существование, распространенность и распределение видов живых организмов в экосистеме определяется тем, может ли уровень одного или нескольких физических или химических факторов быть выше или ниже уровней толерантности этих видов.
Уровень толерантности отдельного организма зависит от его возраста, здоровья, физиологического состояния, генотипа (например, толерантность к спиртному). К постепенно изменяющимся условиям можно адаптироваться (привыкнуть).
Гомеостаз – способность биологического объекта к саморегуляции при изменении условий окружающей среды; для организма сохранение постоянства внутренней среды организма и устойчивость основных физиологических функций при изменении внешних условий.
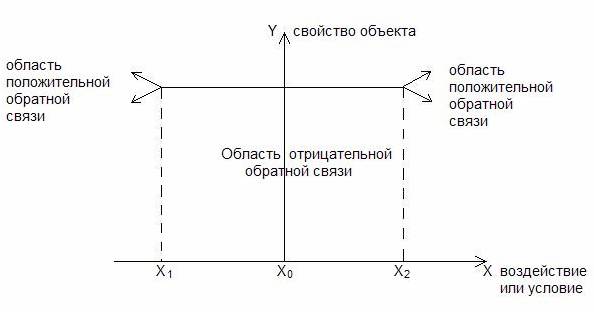
Х1 и Х2 – пороговые значения
Пороговый эффект – малое изменение или воздействие может оказаться критическим и вызвать негативные последствия (если система находится в предпороговой области).
Например, массовая гибель деревьев после длительного воздействия загрязненного воздуха.
Принцип лимитирующего фактора – избыток или недостаток одного абиотического фактора может повлечь за собой ограничение или остановку роста численности популяции в экосистеме, даже если значения других факторов оптимальны.
Лимитирующий фактор – любой фактор, тормозящий рост популяции в экосистеме.
Лимитирующие факторы для наземных экосистем:
- температура;
- вода;
- свет;
- питательные вещества в почве.
Лимитирующие факторы для водных экосистем:
- температура;
- солнечный свет;
- содержание растворенного кислорода;
- соленость.
ПОЧВЕННАЯ МИКРОБИОЛОГИЯ.
Почва представляет освещаемую твердую поверхность Земли, покрытую растительным покровом, и, начиная с силура, примерно 350-400 млн лет назад, обеспечивает выход растений в аэротоп - слой атмосферы между верхушками растений и поверхностью земли. Как операционное биологическое понятие почву удобнее всего определить как "корнеобитаемый слой земной поверхности". При этом за пределами определения окажутся лишенные высшей растительности участки с водорослевой или моховой растительностью и лишенные органического вещества области первичного почво-образовательного процесса: обнаженные грунты, пески, скалы.
Растение пересекает две среды обитания и становится мощным транспортным агентом для потоков вещества. Растение создает своей корневой системой особую область для развития микроорганизмов. В первую очередь это обусловлено эвапотранспирацией растения, обеспечивающей тягу почвенного раствора корнями. Следующим фактором служат корневые выделения, обеспечивающие рост органотрофных организмов ризосферы. Далее, растение вступает в симбиотические отношения прежде всего с грибами-микоризниками, но также и с симбиотическими бактериями, из которых наибольшее внимание заслужили симбиотические фиксаторы азота. Таким образом, задающим параметром становится экофизиология растительного покрова, характеризующего ландшафт.
Примитивные почвы, лишенные растительного покрова, господствовали на поверхности Земли в докембрии и были представлены наземными циано- и альго-бактериальными матами с вероятным ранним появлением грибов, предварительным условием для которых служило появление кислорода в атмосфере. Переходный период к покрову сосудистых растений могли составлять лишайники и мхи. Вынос фотосинтезирующей поверхности в аэротоп мог произойти лишь при создании жестких структур углеродного скелета, представляющего композитную конструкцию из лигноцеллюлозы, включающей фибриллы целлюлозы, склеенные сетью гемицеллюлозы и армированной лигнином. Лигноцеллюлоза у древесных растений на 1/3 состоит из лигнина и в общем синтезе органического вещества в наземных экосистемах составляет не менее 20 Гт/год. Лигноцеллюлоза не подвергается автолизу при отмирании растения и составляет основную массу поступающего в почву органического вещества.
В почвенно-растительном покрове следует различать фотоавтотрофную наземную часть, дополняемую гетеротрофной подземной корневой частью растения, и органотрофную группировку грибов и бактерий. Фотоавтотрофы осуществляют фотосинтез только в дневные часы и зависят от света. Гетеротрофное дыхание идет круглосуточно. Органотрофы действуют круглосуточно, а также вне вегетационного сезона, разлагая листьевой и корневой опад. Разложение лигноцеллюлозы осуществляется в первую очередь грибами. Отсюда вытекают три трофических маршрута:
Миколитический, Целлюлозолитический, Лигнолитический (вторичная биомасса грибов).
Физические условия существования в почве с пористым пространством между минеральными частицами, недостатком влаги обусловливают преимущества для мицелиального строения микроорганизмов. Мицелий может обволакивать твердые растительные остатки и осуществлять гидролиз в непосредственном контакте с ними. Поэтому почву можно рассматривать как растительно-грибную систему. Бактерии развиваются в почвенной влаге и образуют биопленки на поверхности минеральных и органических частиц.
В почвенной системе можно выделить три подраздела с разными условиями жизни для микробиоты: а) первичные продуценты растения с их корневой системой, населенной консорциумом микроорганизмов, включающим микоризу, бактерии ризосферы; б) микробное сообщество, осуществляющее деструкцию мортмассы, конечным продуктом которого является гумус; в) микробное сообщество, взаимодействующее с минеральной частью, конечным результатом чего является превращение минералов материнской породы в педогенные минералы с наиболее характерной группой глинистых минералов. Между этими подразделами системы существует тесное взаимодействие, обусловленное, например, действием корневых выделений и продуктов разложения мортмассы на минералы, но, тем не менее, каждый из разделов удобнее рассматривать отдельно.
Почвенная влага, в том числе пленочная, является местом обитания бактерий и отчасти грибов, мицелий которых может выходить в газовую фазу. Почвенная вода имеет матричный потенциал, зависящий от ее связывания с поверхностью частиц, и осмотический потенциал, зависящий от растворенных веществ. Почвенная вода рассматривается как гравитационная, капиллярная, гигроскопическая. После удаления гравитационной воды остается вода, определяемая почвенной влагоемкостью. Капиллярная вода имеет водный потенциал от -0,01 до -0,03 МПа и доступна для роста микробов, когда наблюдается их наибольшая активность. Ниже -1,5 МПа начинается завядание растений. При -30 МПа организмы сохраняют лишь 10% активности, измеряемой по скорости разложения органического вещества.
Вода в почве входит в состав почвенного раствора, служащего местообитанием бактерий. Состав почвенного раствора определяется прежде всего карбонат-бикарбонатным равновесием, от которого зависят его рН, а также и растворимость минералов как почво-образующей породы, так и педогенных. Важнее всего поведение ти-поморфных элементов: Са, Fe, Al, Si. Почвенный раствор подвержен испарению при засухе и промывному режиму при дождях, и состав его непостоянен. Поэтому почвенная биота должна приспосабливаться к переменному режиму в отношении осмотического давления. Кроме минеральных компонентов, в почвенный раствор поступают органические вещества из корневой системы растений и продукты микробного разложения. Устойчивую часть составляют коллоиды гуминовых веществ, обладающие комплексообразующим действием. Физико-химическая система почвенного раствора зависит от многих переменных и очень сложна. От нее зависит состояние почвенной биоты, и прежде всего бактерий.
РАСПРЕДЕЛЕНИЕ МИКРООРГАНИЗМОВ В ПОЧВЕ.
Очень важным показателем является так называемый "подстилочный коэффициент" - отношение ежегодного опада к подстилке, который показывает скорость разложения растительных остатков. На севере в тундре он составляет 1: 200, а на юге в субтропиках 200: 1. Продукты разложения грибами опада и ветоши служат субстратом для микофильных бактерий, причем особое значение приобретает разложение фенольных соединений. Отмерший мицелий грибов также служит субстратом для бактерий с актиномицетами как характерной группой, идущей по отмершему мицелию. Разложение остатков в хвойных лесах приводит к формированию темноокрашенных гумусовых вод.
Наибольший интерес представляет ассоциация микроорганизмов с корневой системой живых растений. Корни располагаются в почвенном горизонте, наиболее богатом органическим веществом. Взаимодействие с корневой системой включает три области: 1) область почвы с непосредственным воздействие корней - ризосфера; 2) поверхность корня - ризоплана; 3) ткань корня.
В ризосфере наблюдается действие корневых экссудатов, содержащих разнообразные органические вещества, и корневого опада. Экссудаты содержат широкий набор углеводов, аминокислот, органических кислот. Корневой опад дает лигноцеллюлозу и полисахариды слизи (mucigel), составляющие 80% потери углерода корнем. Из суточной ассимиляции 11% уходит на микробное дыхание и 2% -в органическое вещество почвы.
Разнообразие микроорганизмов вблизи корня определяется разнообразием поступающих веществ, разнообразием трофических взаимодействий микроорганизмов между собой (гидролитики и дис-сипотрофы, бактериолитическая петля) и влиянием специфических веществ растений. Здесь можно ожидать очень широкий круг орга-нотрофных аэробных организмов, пищевые потребности которых ориентированы на экссудаты. Как правило, эта область развития микроорганизмов характеризуется избытком органического углерода и лимитирующим содержанием азота и фосфора.
На поверхности листьев в филлосфере развиваются организмы, специфически взаимодействующие с растением, а также паразиты -бактериальные и грибные. Сапрофиты используют гуттацию, слизетечение.