 2013-12-28
2013-12-28 871
871Ранний палеозой: "выход жизни на сушу". Появление почв и почвообразователей. Высшие растения и их средообразующая роль. Тетраподизация кистеперых рыб.
Кембрий: "скелетная революция" и пеллетный транспорт. Эволюция морской экосистемы: кембрий, палеозой и современность.
Итак, мы с вами расстались с вендскими экосистемами в момент, когда они пребывали в состоянии глубочайшего глобального кризиса (глава 6). "Эдиакарский эксперимент" окончился неудачей, и все эти загадочные крупноразмерные существа исчезли с лица Земли, не оставив после себя прямых потомков. Фитопланктонный кризис погубил все огромное разнообразие тогдашних акритарх; уцелели лишь немногие, причем морфологически однотипные, формы. В необратимый упадок пришли и бентосные сообщества прокариот-строматолитообразователей; отныне они будут занимать в биосфере подчиненное положение. Хотя тогда появились и некоторые новые группы животных (например, сабелидитиды, строившие органические трубки), конец венда - это время, когда суммарное разнообразие живых организмов на нашей планете падает до рекордно низкого уровня. И подобно тому, как на развалинах одряхлевшей империи возникает государство молодых энергичных "варваров", на смену докембрийской, прокариотной в своей основе, биосфере стремительно приходит современная, фанерозойская.
Самое же интересное заключается в том, что составляющие эту современную биоту таксоны (а их состав на уровне типов с той поры уже принципиально не менялся!) одновременно обзаводятся минеральным скелетом, причем процесс этот охватывает самые различные группы не только животных, но и водорослей. Вот что пишет, например, один из ведущих специалистов по этому отрезку геологической истории А.Ю. Розанов: "Начнем с того, что поражает любого исследователя, когда он изучает разрезы пограничных отложений докембрия и кембрия. Двигаясь вверх по разрезу от докембрийских пород к кембрийским, мы вдруг обнаруживаем в какой-то момент, что порода насыщена многочисленными и разнообразными остатками организмов, облик которых уже более или менее привычен для нас. Здесь начинают встречаться обычные для всего фанерозоя остатки губок, моллюсков, брахиопод и других организмов. Непривычны только их размеры (первые миллиметры)."
Пытаясь разобраться в причинах этого события, называемого обычно "кембрийской скелетной революцией", надо иметь в виду следующее. Обзавестись твердым скелетом - дело, конечно, "прибыльное", но весьма "капиталоемкое", требующее от организмов больших "первоначальных вложений" по энергии и веществу. Поэтому строить скелет способны лишь существа с достаточно высоким уровнем энергетического обмена, каковой возможен лишь начиная с некоторого уровня содержания на планете свободного кислорода (о "гипотезе кислородного контроля" - см. главу 5). Проанализируем теперь под этим углом зрения докембрийскую экологическую ситуацию.
Начать тут придется издалека. Одной из самых важных задач современной океанологии является детальное изучение морских течений; это, помимо прочего, весьма важно для практики судовождения. С этой целью в воды Мирового океана ежегодно бросают, точно зафиксировав место и время, многие тысячи специальных буев; выловив впоследствии такой буй в другой точке океана, мы получаем представление о направлении и скорости движения переносивших его водных масс. Исследования эти стоят больших денег, а поскольку подавляющее большинство буев при этом безвозвратно теряется, эффективность метода не слишком высока. В связи с этим встал вопрос: а нельзя ли вместо буев найти какой-нибудь естественную "метку" для океанских водных масс?
Такая "метка" действительно существует: это просто-напросто... пыль, которая всегда содержится в атмосфере и, понятное дело, постоянно оседает не только на крышку вашей парты, но и на поверхность океана. Химический состав этой атмосферной пыли в каждой точке Земли свой - на него влияет геохимия соответствующей территории, тип вулканической активности и т.д. А поскольку возможности аналитической химии по нынешнему времени почти безграничны (криминалисты, например, с легкостью определяют, в каком из месторождений мира было добыто контрабандное золото), то для реконструкции картины течений кажется достаточным просто зачерпнуть толику морской воды и установить, в каких именно местах Земли выпадала содержащаяся в воде пыль. Океанологи с энтузиазмом принялись за работу, и спустя небольшое время с изумлением обнаружили, что атмосферной пыли (равно как и вообще взвеси) в океанской воде практически не содержится; на подоконнике, к примеру, пыль будет скапливаться до тех пор, пока ее не сотрет дежурный по классу, а вот из океана она странным образом куда-то улетучивается. Быстро осесть на дно пылинка не может - это можно доказать простыми физическими расчетами. В чем же дело?
К тому времени было уже хорошо известно, какую титаническую работу осуществляют в морях животные-фильтраторы, и прежде всего двустворчатые моллюски; согласно расчетам, одни только уже закартированные устричные банки прогоняют сквозь себя объем воды, равный всему Мировому океану, примерно за месяц! Однако двустворки очищают именно придонные слои воды, куда, как мы помним, пылинкам попасть довольно сложно... Вот тогда-то и выяснилось, что планктонные фильтраторы, и прежде всего - ракообразные, играют в очищении океана роль ничуть не меньшую, чем бентосные. Отсутствие в водах океана сколь-нибудь ощутимых количеств пыли, которая постоянно оседает на его поверхность, является результатом деятельности планктонных ракообразных: они очень быстро отфильтровывают взвесь, упаковывают ее в формируемые их пищеварительным трактом компактные комки отработанной органики - фекальные пеллеты, и отправляют на дно. Без пеллетной транспортировки взвеси мутность воды в океанах была бы несравненно выше.
Именно такая ситуация и должна была наблюдаться в далеком прошлом. Как мы с вами помним, отсутствие в докембрии наземной растительности приводило к многократному усилению эрозии и формированию специфических выположенных ландшафтов - "не суша-не море" (глава 5). Сток с континентов был в основном "плащевым" (т.е. по всей протяженности береговой линии), а равнинные реки и озера, являющиеся ныне главными отстойниками взвеси, отсутствовали. Воды, стекавшие с суши, были мутными, причем самой замутненной в итоге оказывалась именно прибрежная часть океана, наиболее богатая биогенами. Это чрезвычайно затрудняло утилизацию биогенов фитопланктоном из-за узости фотической зоны и резко ограничивало общую продуктивность планктонной экосистемы.
С другой стороны, иной оказывалась и организация самой водной массы. В современных океанах имеется верхний слой воды с резко выраженными сезонными колебаниями температуры - эпиталасса, и нижний, где температура на протяжении всего года практически неизменна - гипоталасса (в озерах им соответствуют эпи- и гиполимнион). На границе между этими слоями располагается узкая зона резкого изменения температуры - термоклин; в современных морях он находится на глубинах от 15 до 100 м. В условиях же замутненной воды энергия солнца практически не проникает глубже первых метров, и термоклин оказывается "подтянутым" под самую поверхность; водная масса оказывается жестко стратифицированой - т.е. разделенную на почти неперемешивающиеся между собою слои: узкую перегретую эпиталассу и относительно холодную, но при этом практически лишенную кислорода гипоталассу. Для этого времени были обычны донные осадки, резко обогащенные неокисленной органикой ("черные сланцы"), которые считают несомненным показателем придонной аноксии (бескислородных условий); позже (начиная с кембрия) их распространение резко сократится. Ситуация аноксии ("заморные обстановки") сильно затрудняла, или даже вообще исключала, существование донной фауны.
И вот на границе венда и кембрия произошло событие, важность которого палеонтологи осознали лишь в самое последнее время (впервые эту идею высказал в 1985 г. А.Г.Пономаренко, а теперь она подтверждена геохимическими данными): в составе докембрийского зоопланктона (бесскелетного, и потому не имевшего шансов захорониться - см. главу 6) возникли достаточно эффективные фильтраторы, формировавшие фекальные пеллеты. Единичные фекальные пелеты были известны и раньше (еще в протерозое), однако именно начало кембрия - это время массового их распространения в морских отложениях.
Следствием этого, вроде бы малозначительного, события стал целый каскад экосистемных перестроек. Уменьшение мутности воды привело к резкому расширению фотической зоны и росту продуктивности экосистем. Расширение же эпиталассы привело к разрушению жесткой температурной стратификации и некоторому обогащению кислородом нижележащих водных слоев. Мало того; мы с вами помним (из главы 6), что именно дисперсная органика, опускающаяся из фотической зоны, является причиной аноксии в придонных слоях - ибо на ее окисление там расходуется кислород; многие исследователи даже употребляют термин "эвтрофный океан". Теперь же органика эта стала, вместе с неорганическими частичками, поступать на морское дно в концентрированном, "упакованном" виде - в пеллетах. Органический детрит, обогащающий донные осадки, - это уже не удушающий эвтрофикатор, а ценный пищевой ресурс; содержание же кислорода в придонных слоях в результате возрастает до уровня, допускающего существование макроскопической донной фауны. Начиная с этого момента в системе возникает положительная обратная связь: придонная фауна в свою очередь начинает очищать воду от органической и минеральной взвеси, количество кислорода в итоге опять возрастает, это позволяет еще дальше наращивать разнообразие живых организмов, и т.д.
Количество свободного кислорода на планете тем временем тоже увеличивается. Во второй половине венда в уже знакомых нам амфибиотических ландшафтах ("ни суша, ни море") место цианобактериальных матов, с их потрясающим по совершенству балансом в производстве и потреблении органики, занимают сообшества водорослей (главным образом - нитчатых), которые иногда называют "водорослевыми болотами". В этих болотах, чрезвычайно широко распространенных во внетропической зоне, захоранивается огромное количество неокисленной органики, а именно такая ситуация (как мы помним из главы 5) вызывает накопление свободного кислорода. По-видимому, сыграло свою роль и крупное Бойканурское оледенение, маркирующее границу венда и кембрия: мы уже помним (см. главу 6), что наличие льдов и психросферы облегчает "закачку" кислорода в глубокие слои океана, вентиляция же дна благоприятствует развитию бентоса.
И вот, наконец, содержание свободного кислорода в океанах достигает той пороговой отметки, начиная с которой становится "экономически оправданным" обзаводиться такой дорогостоящей "высокой технологией", как минеральный скелет. Эта возможность немедленно и одновременно реализуется самыми разными группами животных - от простейших (радиолярии [21] и форамениферы) и губок до членистоногих и моллюсков, - а также растений (известковые водоросли): начинается "скелетная революция" в собственном смысле. Важная деталь: как отмечает А.Ю. Розанов (1986), химический состав этих скелетов, возникающих в самом начале кембрия, настолько разнообразен, что почти полностью исчерпывает все те варианты, которые будут затем возникать на протяжении всего фанерозоя.
Самым лучшим, уникальным, местонахождением кембрийской фауны считают среднекембрийские сланцы Бёджес, открытые в 1912 г. в канадских Скалистых Горах (богатейшие местонахождения в Восточной Сибири, Китае и Гренландии изучены пока недостаточно); тип сохранности в Бёджес таков, что там могут захораниваться и "мягкие" организмы, не имевшие скелета. Так вот, оказалось, что число таких бесскелетных форм в кембрийской фауне даже больше, чем в вендской (хотя доля их теперь невелика). Это служит сильным доводом в пользу того, что в кембрии не появляется вдруг, как по мановению волшебной палочки, некая принципиально новая - "скелетная" - фауна (так иногда полагали прежде), а одевается скелетом та фауна, что уже существует - едва лишь открывается для этого возможность.
Другой неожиданный вывод следует из изучения открытых в конце восьмидесятых годов в Швеции среднекембрийских фосфатных конкреций Ёрстен. При тамошнем типе сохранности фиксируются не покровные ткани (как в Бёждес), а микроскопические щетинки и иные твердые покровные элементы, видимые лишь под сканирующим электронным микроскопом. Оказалось, что в то время существовал многочисленный и очень разнообразный микрозоопланктон - трохофоры (личинки кольчатых червей), зоеа (личинки ракообразных) и т.п. Это позволяет вновь вернуться к гипотезе А.Ю.Розанова (1986) о том, что в кембрии в действительности произошло лишь увеличение размеров уже существовавших на тот момент планктонных организмов, которые по этой причине "выпали в осадок" - перешли к бентическому образу жизни; дополнительные аргументы обретают и построения М.Б. Бурзина (см. выше) о эволюции планктонных экосистем.
Надо заметить, что мир первых скелетных организмов, сложившийся в начале кембрия (немакит-далдынский ярус), был крайне своеобразен и в чистом виде просуществовал очень недолго, лишь до середины этого периода - хотя отдельные ого представители дожили до середины ордовика. В это время уже существовали достаточно многочисленные животные, имевшие раковины (моллюски и брахиоподы) и жилые трубки (сабелидиты, хиолиты) и т.п.; разнообразнейшие мелкие зубчики - конодонты представляли собою части ротовых органов древнейших вторичноротых - щетинкочелюстных и хордовых. Весьма характерны для этого мира приапулиды (в современных морях эта реликтовая группе насчитывает лишь 20 видов), а также лобоподы (представленные ныне микроскопическими тихоходками и обитающими в подстилке влажных тропических лесов онихофорами); видимо, именно к лобоподам принадлежат многие загадочные кембрийские организмы, вроде галлюциногении (оцените название!). Имелись и и многочисленные артроподоподобные формы (см. ниже) (рисунок 25, а-д).
Основу тогдашних экосистем, однако, составляли водоросли, строившие небольшие известковые постройки - биогермы, и чрезвычайно любопытная, как считалось, вымершая группа организмов - археоциаты. Археоциаты (по-гречески - "древние кубки") представляют собой небольшие прикрепленные ко дну бокалы с двуслойной стенкой (рисунок 24); диаметр их колеблется от нескольких миллиметров до нескольких сантиметров, хотя в Сибири найдены и исполинские формы размером до 1,5 м. Природа их долго вызывала споры (неясно было даже - животные это или растения, или вообще отдельное царство), однако в последнее время почти все исследователи согласны с тем, что их следует относить к губкам. Более того - не так давно в Тихом океане была найдена странная глубоководная губка Vacletia, оказавшаяся по рассмотрении дожившим до наших дней археоциатом и пополнившая собою галерею "живых ископаемых" (вроде кистеперой рыбы или гинкго).
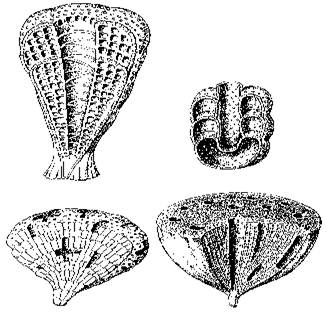 | 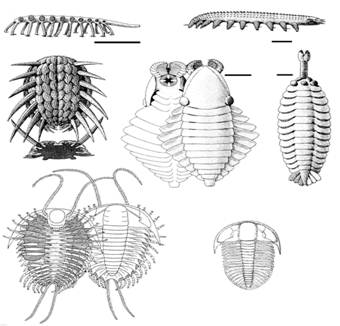 |
| РИСУНОК 24. Археоциаты. (а) - Archaeocyatida, (б) - Capsulocyatida, (в) - Kazachstanicyatida, (г) - Archaeocyatida. | РИСУНОК 25. Различные кембрийские животные: лобоподы (а-б), члнистоногие неясного систематического положения (в-д), трилобиты (е-ж). (а) - Xenusion; (б) - Aysheaia; (в) - Wiwaxia; (г) - Anomalacaris; (д) - Opabinia; (е) - Olenoides, (ж) -Ogygopsis. |
Губки занимают совершенно обособленное положение в системе животного царства и могут быть противопоставлены всем остальным многоклеточным животным: есть веские основания считать, что они ведут свое происхождение от иной группы простейших, нежели прочие метазоа. Существует, например, остроумная и неплохо аргументированная гипотеза Й. Райтнера (1991), согласно которой губки возникают как симбиоз бактериальной колонии с хоанофлагеллятными простейшими [22]. Достигнутый губками уровень организации не позволяет назвать их многоклеточными в строгом смысле ("многотканевыми" по Корлиссу), так что вроде бы нет ничего удивительного в том, что именно эта примитивнейшая группа животных открывает собою фанерозойскую летопись. Но это только на первый взгляд.
Дело в том, что крайне низкий уровень клеточной интеграции оставляет губкам единственно возможную "профессию" - прикрепленный пассивный фильтратор органической взвеси. Любая губка (включая археоциат) представляет собою открытый сверху мешок, стенки которого пронизаны каналами; вода постоянно движется сквозь них во внутреннюю (парагастральную) полость, выходя затем наружу через верхнее отверстие (устье), а входящие в состав стенок каналов хоаноциты (воротничковые жгутиковые клетки) отфильтровывают содержащуюся в воде органику и бактерий. Вопрос для школьной олимпиады: какая сила заставляет воду двигаться сквозь каналы в неподвижной стенке? Ответ: та же самая, что создает печную тягу - разница давлений на высоте поддувала и на высоте конца трубы в соответствии с законом Бернулли [23]: если водная среда, в которой находится губка, имеет ненулевую скорость, то слой движущейся жидкости над устьем создает "подсос". Известное дело: чем выше труба - тем лучше тяга; соответственно, губка может прокачивать сквозь себя воду лишь когда ее устье приподнято над субстратом. (Сходный механизм обеспечивает вентиляцию нор сусликов и иных грызунов. Тот из выходов, через который при рытье норы выбрасывали грунт, оказывается заключен в холмик-"кротовину" и несколько приподнимается над землей; в итоге внутри норы возникает устойчивый ток воздуха от "низкого" выхода к "высокому"). Подъем же устья над субстратом можно обеспечить лишь при наличии твердого опорного скелета - органического или минерального (у современных губок есть оба варианта). Отсутствие в предшествующих отложениях этих легко обнаруживаемых скелетных элементов - спикул - свидельствует о том, что данная группа живых организмов действительно возникла лишь в начале кембрия (возможно, именно по схеме Райтнера - см. выше), т.е. когда уже несомненно существовали и настоящие многоклеточные животные.
Вся геологическая история археоцат (появление, расцвет - около 300 родов, - упадок и вымирание) укладывается в крохотный по геологическим меркам интервал в 15-20 млн. лет - случай совершенно уникальный. Уже во второй половине кембрия эта процветавшая в начале периода группа исчезает "как с белых яблонь дым", и при этом на смену ей не приходит никто - то есть конкурентное вытеснение здесь предположить трудно. Создается впечатление, что археоциаты эксплуатировали некий ресурс, который сперва был в изобилии, а затем стал резко дефицитным. Сопоставляя экологию археоциат (по аналогии с современными губками) и изложенную выше картину вендско-кембрийской биосферной перестройки можно предположить следующее. Группа эта процветала в тот относительно краткий и принципиально неустойчивый период, когда развитие пеллетного транспорта уже обогатило придонные слои кислородом, но еще не сконцентрировало большую часть органики внутри осадка; этот расклад, как полагает А.Г. Пономаренко, должен быть весьма благоприятен для "тонких" фильтраторов. Для того, чтобы эксплуатировать открывшийся ресурс, нет нужды в "квалифицированной рабочей силе" - вполне достаточно макроскопических организмов, даже не являющихся многотканевыми. Однако дальнейшее развитие ситуации с запасанием органики в осадке (что позволяет и сделать ее переработку более равномерной, и улучшить снабжение кислородом придонных слоев) резко ухудшает положение бактерий и пассивных фильтраторов, но зато благоприятствует илоедам. Время археоциат кончается, и наступает время трилобитов (рисунок 25, е-ж) [24].
Давайте теперь вернемся к графику, показывающему изменения разнообразия фауны на протяжении фанерозоя (Рисунок 23). Обсуждаемые нами события знаменуют собой формирование новой биоты, которая сохраняет достаточное единство на протяжении всего палеозоя, и гораздо более разнообразна, чем прежняя, "кембрийская" - и в таксономическом, и в экологическом отношении. В современной экологии достаточнно строго установлено, что смена эвтрофных условий на олиготрофные вызывает в экосистеме более "мелкую нарезку" экологических ниш, и соответственно, рост биологического разнообразия. Все это достаточно хорошо соответствует изложенным выше представлениям об "олиготрофизации" кембрийского океана за счет пеллетного транспорта.
В бентосе тем временем к обитателям раковин и жилых трубок добавляются существа с принципиально иными типами твердого скелета - внутренним (хордовые) и подвижным наружным (членистоногие). Ситуация с членистоногими интересна тем, что они появились позднее многих типов животного царства (во втором веке кембрия - атдабанском), однако сразу в составе практически всех своих подтипов и классов - и ракообразные, и хелицеровые, и вымершие трилобиты, и множество удивительных существ, точная систематическая принадлежность которых до сих пор неясна (рисунок 25, в-д).
Недавно стало выясняться, что ситуация с кембрийскими "членистоногими" (Arthropoda) сходна с той, что была с вендобионтами (см. главу 5): по мере изучения оказывается, что значительная часть из них не может быть отнесена к настоящим артроподам. Например, выяснилось, что у аномалакариса - самого крупного, к слову сказать, животного того времени, достигавшего в размере 70 см - ротовые части состоят из четырех концентрически сходящихся лепестков, образующих нечто вроде зажимов цангового карандаша (рисунок 25, г). Эти лепестки никоим образом не могут представлять собою видоизмененные конечности, что является одним из ключевых диагностических признаков типа членистоногих.
По всей видимости, целый ряд неродственных между собой групп начинает в это время независимо приобретать отдельные признаки членистоногих, и в результате они независимого достигают артроподного уровня организации. Подробнее о процессах параллельной эволюции, подобных этой "артроподизации", мы поговорим позже (в главе 12), пока же для нас существенно лишь само возникновение принципиально нового типа скелета, обладатели которого начинают играть ключевую роль в формирующихся экосистемах. Так, трилобиты становятся, по-видимому, основными потребителями того самого "упакованного" органического детрита, обогащающего донные осадки, и могут считаться настоящей эмблемой палеозоя.
К середине кембрия продуктивность и биологическое разнообразие достигают такого уровня, что у экосистем возникает нужда в "управляющем блоке" - верхних трофических уровнях, представленных специализированными хищниками-макрофагами. И вот начинается интереснейший процесс - борьба за этот "государственный военный заказ" между несколькими, самыми мощными, "корпорациями" - наиболее эволюционно продвинутыми группами животных. Из курса зоологии вам должно быть известно, что самыми высокоорганизованными среди первичноротых являются членистоногие и моллюски, а вершину эволюции вторичноротых представляют собой позвоночные. Именно эти группы и начинают соревнование за открывшуюся вакансию.
Первыми успеха добились членистоногие и близкие к ним артроподоподобные формы. Самым крупным хищником тех времен был аномалокарис; хищниками, как считают сейчас многие исследователи, были и самые первые трилобиты - лишь позднее эта группа перешла к детритофагии [25]. Однако вскоре вперед вырвались моллюски: появившиеся в конце кембрия головоногие (это были малоподвижные формы, родственные современному наутилусу, но с незакрученной конической раковиной, так называемые прямые головоногие) безраздельно господствовали в придонных слоях океана примерно до середины силура; на ордовик приходится пик видового разнообразия этой группы животных (около 150 родов). Хищники из числа членистоногих (ракообразные и водные хелицеровые - мечехвосты и ракоскорпионы) и позвоночных (бесчелюстные - родственики современных миног) оказываются "неконкурентоспособными" и занимают в экосистемах подчиненное положение.
Положение меняется в силуре, когда среди членистоногих появились крупноразмерные ракоскорпионы, а среди позвоночных - первые челюстноротые (настоящие рыбы). Судя по всему, обе эти группы возникли в опресненных прибрежных акваториях (типа лагун): здесь они были на первых порах избавлены от конкуренции с головоногими, которые избегают участков моря, "отравленных" стекающими с суши пресными водами. Именно в это время ракоскорпионы (например, Pterygotus) достигают длины свыше 2 м (а если с клешнями - то почти 3) и становятся властелинами силурийских морей - по крайней мере, их прибрежных участков (рисунок 26). Прямые головоногие не выдерживают конкуренции, и их разнообразие заметно сокращается; попытки спастись за счет гигантизма (раковины Endoceras, например, достигали 4 м в длину - хотя сам моллюск был, понятное дело, много меньше) не имели успеха.
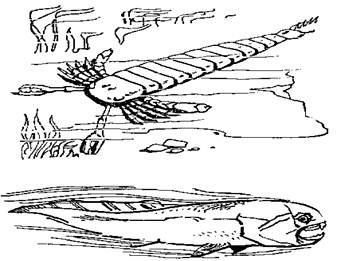 |
| РИСУНОК 26. Самые страшные хищники раннего палеозоя - ракоскорпион и динихтис. |
Однако торжество членистоногих (в лице ракоскорпионов) оказалось недолгим, и виною тому была третья "конкурирующая корпорация" - позвоночные. До сих пор те держались в тени, и использовали лишь стратегию пассивной защиты - создание мощной наружной "брони"; таковы были панцирные бесчелюстные - остракодермы, и панцирные рыбы - плакодермы. И вот в девоне одна из групп плакодерм - артродиры - отказалась от "бронирования корпуса" в обмен на скорость и перешла к активному хищничеству. Вот тогда-то и проявились все те - до поры скрытые - преимущества, что возникли ранее в результате превращения первой пары жаберных дуг в подвижные челюсти - великолепное средство нападения и защиты. Существа эти могли увеличивать размеры тела практически неограниченно: такие артродиры, как динихтис достигали 10 м в длину - одни из самых крупных рыб за всю историю (рисунок 26), тогда как ракоскорпионы уже "уперлись" в физиологический предел размеров для членистоногих. В итоге артродиры стали для того времени фактически "абсолютным оружием", превосходя любого противника и по вооружению, и по скорости.
Членистоногие проиграли позвоночным эту схватку за крупный размерный класс вчистую: ничего подобного двухметровым силурийско-раннедевонским ракоскорпионам среди них не появится уже никогда. Отныне они будут методично осваивать малый и средний размерный классы (длина тела самых крупных членистоногих - камчатского краба и мечехвоста - не превышает 70 см), в чем и преуспеют: это, как-никак, самый процветающий ныне тип животного царства. А вот головоногие сумели тогда дать позвоночным достойный эволюционный ответ: хотя численность придонных прямых головоногих продолжала сокращаться, суммарное разнообразие группы начиная с середины девона (эмский век) стремительно пошло вверх. К концу периода (фаменский век) оно достигло своего максимума (почти 200 родов) благодаря появлению свернутых головоногих, имеющих спирально закрученную раковину - аммонитов.
Дело в том, что прямые наутилоидеи обладали достаточно совершенным аппаратом регулирования плавучести (они меняли заполнение водою пустых камер раковины - в точности как подводная лодка заполняет и опорожняет балластные цистерны), но вот с горизонтальными перемещениями у них были большие проблемы. Головоногие движутся реактивным способом, выталкивая воду из мантийной полости через воронку, направленную вперед и располагающуюся в районе головы. Первые наутилоидеи "висели" вниз головой в придонных слоях воды и могли лишь "падать" на свою добычу сверху (рисунок 27, а). В дальнейшем они "завалили" раковину, сместив центр ее тяжести за счет неравномерного отложения карбоната кальция (на брюшной стороне больше, чем на спинной); вся конструкция в результате обрела способность к продольному движению (рисунок 27, б). Однако эта конструкция принципиально неустойчива в движении, т.к. центр тяжести и точка приложения реактивной тяги (воронка) должны находиться на противоположных концах тела животного: малейшее отклонение от линии тяги - и движущийся моллюск начнет кувыркаться. А вот у свернутых головоногих центр тяжести их спиральной раковины почти совпадает с сифоном, что и обеспечивает устойчивое продольное движение. Именно поэтому свернутые головоногие стали первой группой хищников, освоивших толщу воды. (Здесь необходимо заметить, что рыбы, например, будут продолжать "ходить по дну" почти до середины мезозоя: лишь тогда костные рыбы "изобретут" плавательный пузырь, а среди хрящевых рыб - появятся акулы современного типа, которые "не тонут" из-за того, что находятся в постоянном движении.) Видимо, именно освоение трехмерного пространства привело к возникновению у позднепалеозойских головоногих самых совершенных (среди беспозвоночных) глаз и чрезвычайно усложненного мозга - то же самое, что произойдет позднее с птицами.
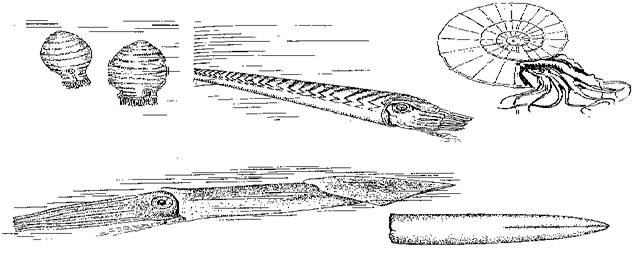
РИСУНОК 27. Палеозойские и мезозойские головоногие: (а) - исходная жизненная форма головоногих (архаичная наутилоидея Mandaloceras); (б) - прямое головоногое (наутилоидея Michelinoceras); (в) - свернутое головоногое (аммонит); (г) - внутренераковинное головоногое (белемнит и его раковинвина - "чертов палец").
Головоногие будут и дальше, на протяжении всего мезозоя, формировать верхние трофические уровни морских экосистем, успешно конкурируя не только с рыбами, но и с морскими рептилиями (ихтиозаврами, плезиозаврами и пр. - см. далее, главу 10). Многие из этих рептилий сами охотились на головоногих (в раковинах крупных аммонитов иногда находят застрявшие зубы этих хищников), а те в основном использовали стратегию пассивной защиты - увеличивая размеры тела. К юре аммониты достигли максимального своего размера, однако с появлением короткошейных плезиозавров - плиозавров, черепа которых достигали 3 м в длину, - эта стратегия себя исчерпала. Аммониты начали мельчать, пытаясь уйти за нижний предел оптимального размера жертвы (см. главу 6-а) [26], а разнообразие их - уменьшаться.
И вновь головоногие нашли выход: с этого времени начинается расцвет групп, имеющих внутреннюю раковину - белемнитов и происходящих от них кальмаров. Вообще-то белемниты появились еще в карбоне, однако до сих пор они пребывали на вторых ролях. Теперь же, когда возможности пассивной защиты оказались исчерпаны, стратегия белемнитов - отказ от раковины в обмен на увеличение скорости передвижения - оказывается генеральной линией эволюции головоногих. Судя по всему, внутреннераковинные головоногие оказались самыми скоростными обитателями мезозойского океана; они, казалось бы, должны столкнуться с тою же проблемой устойчивости в движении, что и прямые головогоногие (см. выше), однако их тело, лишенное раковины, способно выравнивать "полет" при помощи плавников-стабилизаторов (рисунок 27, г). При этом отсутствие таких ограничителей роста, как твердые покровы членистоногих, позволяет им достигать очень крупных размеров. Современный гигантский кальмар достигает в длину 18 м (со щупальцами) и является самым крупным беспозвоночным (на коже китов находили отпечатки присосок кальмаров, длина которых оценивается в 30 м), а ведь в прошлом вполне могли существовать и более крупные виды! Как бы то ни было, внутреннераковинные головоногие успешно дожили до наших дней и вымирать явно не собираются. Достаточно сказать, что в пробах грунта из глубоководных частей Тихого океана число челюстей кальмаров на один квадратный метр варьирует от нескольких десятков до нескольких сотен, что дает некоторое представление о численности (и экологической роли) этих животных в современных морях. Изменения в сообществе водных позвоночных мы подробнее проанализируем чуть позже, в главе 11. Здесь же мы лишь отметим, что с начала триаса верхние трофические уровни морских экосистем оказываются совершенно закрытыми для рыб: их формируют недавние вселенцы в море - хищные рептилии. Однако во второй половине мезозоя (примерно в одно время с началом расцвета внутреннераковинных головоногих) рыбы возвращают себе лидирующие позиции: в морях появились акулы современного типа - с челюстью, подвижной относительно мозговой капсулы, что позволяет им отрывать куски от жертвы большого размера (например, кита), а не глотать ее целиком [27]. Именно они, возможно, конкурентно вытеснили по крайней мере часть морских рептилий (например, ихтиозавров). Самые крупные акулы жили в миоцене: Carcharodon megalodon (близкая родственница знаменитой белой акулы) достигала в длину 30 м; питалось это чудовище, по всей видимости, небольшими китами, пик разнообразия которых приходится как раз на время ее существования (рисунок 28). Самые же крупные из современных хищных акул [28] (белая, тигровая) питаются в значительной степени другими акулами, являясь, таким образом, подлиным венцом трофической пирамиды.
 |
| РИСУНОК 28. Челюсти ископаемой акулы Carcharodon megalodon. |
Вплоть до самого недавнего времени человек выносил из школьного учебника биологии и популярных книг по теории эволюции такую примерно картину события, именуемого обычно "Выходом жизни на сушу". В начале девонского периода (или в конце силурийского) на берегах морей (точнее - морских лагун) появились заросли первых наземных растений - псилофитов (рисунок 29, а), положение коих в системе растительного царства остается не вполне ясным. Растительность сделала возможным появление на суше беспозвоночных животных - многоножек, паукообразных и насекомых; беспозвоночные, в свою очередь, создали пищевую базу для наземных позвоночных - первых амфибий (ведущих свое происхождение от кистеперых рыб) - таких, как ихтиостега (рисунок 29, б). Наземная жизнь в те времена занимала лишь чрезвычайно узкую прибрежную полоску, за которой простирались необозримые пространства абсолютно безжизненных первичных пустынь.
 |
| РИСУНОК 29. (а) - Реконструкция ландшафта Райни (девон Шотландии) с зарослями псилофитов; (б) - древнейшее наземное позвоночное - ихтиостега (девон Гренландии). |
Так вот, согласно современным представлениям, в означенной картине неверным (или, по меньшей мере, неточным) является практически всё - начиная с того, что достаточно развитая наземная жизнь достоверно существовала много раньше (уже в следующем за кембрием ордовикском периоде), и кончая тем, что упомянутые "первые амфибии" наверняка были существами чисто водными, не имеющими связи с сушей. Дело, однако, даже не в этих частностях (о них мы поговорим в свой черед). Важнее другое: скорее всего, принципиально неверна сама формулировка - "Выход живых организмов на сушу". Есть серьезные основания полагать, что сухопутные ландшафты современного облика в те времена вообще отсутствовали, и живые организмы не просто вышли на сушу, а в некотором смысле создали ее как таковую. Впрочем, давайте по порядку.
Итак, первый вопрос - когда; когда же все-таки на Земле появились первые несомненно наземные организмы и экосистемы? Однако тут сразу возникает встречный вопрос: а как определить, что некий вымерший организм, с которым мы столкнулись - именно наземный? Это вовсе не так просто, как кажется на первый взгляд, ибо принцип актуализма здесь будет работать с серьезными сбоями. Типичный пример: начиная с середины силурийского периода в палеонтологической летописи появляются скорпионы - животные по нынешним временам вроде бы сугубо сухопутные. Однако сейчас уже достаточно твердо установлено, что палеозойские скорпионы [29] дышали жабрами и вели водный (или, по крайней мере, амфибиотический) образ жизни; наземные же представители отряда, у которых жабры превращены в характерные для паукообразных легкие "книжечного" типа (book-lungs), появились лишь в начале мезозоя. Следовательно, находки в силурийских отложениях скорпионов сами по себе ничего (в интересующем нас плане) не доказывают.
Более продуктивно здесь, как кажется, отслеживать появление в летописи не наземных (по нынешним временам) групп животных и растений, а определенных анатомических признаков "сухопутности". Так, например, растительная кутикула с устьицами и остатки проводящих тканей - трахеид наверняка должны принадлежать наземным растениям: под водой, как легко догадаться, и устьица, и проводящие сосуды ни к чему... Однако существует и иной - воистину замечательный! - интегральный показатель существования в данное время наземной жизни. Подобно тому, как показателем существования на планете фотосинтезирующих организмов является свободный кислород, показателем существования наземных экосистем может служить почва: процесс почвообразования идет только на суше, а ископаемые почвы (палеопочвы) хорошо отличимы по структуре от любых типов донных отложений.
Надо заметить, что почва сохраняется в ископаемом состоянии не слишком часто; лишь в последние десятилетия на палеопочвы перестали смотреть как на некую экзотическую диковинку и начали их систематическое изучение. В итоге в изучении древних кор выветривания (а почва есть не что иное, как биогенная кора выветривания) произошла подлинная революция, буквально перевернувшая прежние представления о жизни на суше. Самые древние палеопочвы найдены в глубоком докембрии - в раннем протерозое; в одной из них, имеющей возраст 2,4 млрд лет, С. Кемпбелл (1985) обнаружил несомненные следы жизнедеятельности фотосинтезирующих организмов - углерод со смещенным изотопным отношением 12С/13С. В этой связи можно упомянуть и обнаруженные недавно остатки цианобактериальных построек в протерозойских карстовых полостях: процессы карстования - образование котловин и пещер в водорастворимых осадочных породах (известняки, гипсы) - могут идти только на суше.
Другим фундаментальным открытием в этой области следует считать обнаружение Г. Реталляком (1985) в ордовикских палеопочвах вертикальных норок, прорытых какими-то достаточно крупными животными - видимо, членистоногими или олигохетами (дождевыми червями) [30]; в этих почвах нет никаких корней (которые обычно сохраняются очень хорошо), но есть своеобразные трубчатые тельца - Реталляк интерпретирует их как остатки несосудистых растений и/или наземных зеленых водорослей. В несколько более поздних, силурийских, палеопочвах найдены копролиты (окаменевшие экскременты) каких-то почвообитающих животных; пищей им, судя по всему, служили гифы грибов, составляющие заметную долю вещества копролитов (впрочем, не исключено, что грибы могли и вторично развиться на органике, содержащейся в копролитах).
Итак, к настоящему моменту два факта могут считаться установленными достаточно твердо:
1. Жизнь появилась на суше очень давно, в среднем докембрии. Она была представлена, судя по всему, различными вариантами водорослевых корок (в том числе - амфибиотическими матами) и, возможно, лишайниками [31]; все они могли осуществлять процессы архаичного почвообразования.
2. Животные (беспозвоночные) существовали на суше по меньшей мере с ордовика, т.е. задолго до появления высшей растительности (чьи достоверные следы по-прежнему остаются неизвестными до позднего силура). Средой обитания и пищей этим беспозвоночным могли служить упомянутые выше водорослевые корки; при этом сами животные неизбежно становились мощным почвообразующим фактором.
Последнее обстоятельство заставляет вспомнить одну старую дискуссию - о двух возможных путях заселения суши беспозвоночными. Дело в том, что неморские ископаемые этого возраста были очень редки, и все гипотезы на сей предмет казались лишь более или менее убедительными спекуляциями, не подлежащими реальной проверке. Одни исследователи предполагали, что животные вышли из моря напрямую - через литораль с водорослевыми выбросами и иными укрытиями; другие настаивали на том, что сперва были заселены пресноводные водоемы, и лишь с этого "плацдарма" началось впоследствии "наступление" на сушу. Среди сторонников первой точки зрения выделялись своей убедительностью построения М.С. Гилярова (1947), который, основываясь на сравнительном анализе адаптаций современных почвообитающих животных, доказывал, что именно почва должна была служить первичной средой обитания самых ранних жителей суши [32]. При этом надо учитывать, что почвенная фауна действительно крайне плохо попадает в палеонтологическую летопись и отсутствие ископаемых "документов" здесь вполне объяснимо. У этих построений, однако, был один по-настоящему уязвимый пункт: а откуда же взялась сама эта почва, если в те времена еще не было наземной растительности? Всем ведь известно, что почвообразование идет при участии высших растений - сам Гиляров называл настоящими почвами лишь те, что связаны с ризосферой, а все прочее - корами выветривания... Однако теперь - когда стало известно, что примитивное почвообразование возможно и с участием одних лишь низших растений - концепция Гилярова обрела "второе дыхание", а недавно была впрямую подтверждена данными Реталляка по ордовикским палеопочвам.
С другой стороны, несомненные пресноводные фауны (которые содержат, помимо всего прочего, дорожки следов на поверхности осадка) появляются намного позже - в девоне. В их состав входят скорпионы, мелкие (примерно в ладонь величиной) ракоскорпионы, рыбы и первые неморские моллюски; среди моллюсков есть и двустворчатые - длительно живущие организмы, неспособные переносить заморы и пересыхание водоемов. Фауны же с такими бесспорно-почвенными животными, как тригонотарбы ("панцирные пауки") и растительноядные двупарноногие многоножки существуют уже в силуре (лудловский век). А поскольку водная фауна всегда попадает в захоронения на порядок лучше, чем наземная, то все это позволяет сделать еще один вывод:
3. Почвенная фауна появилась существенно раньше пресноводной. То есть - по крайней мере для животных пресные воды не могли играть роль "плацдарма" при завоевании суши.
Этот вывод, однако, заставляет нас вернуться к тому самому вопросу, с которого мы начинали свои рассуждения, а именно: вышли живые организмы на сушу или фактически создали ее как таковую? А.Г. Пономаренко (1993) полагает, что все сообщества, о которых шла речь выше, на самом деле трудно с определенностью назвать "наземными" или "сообществами внутриконтинентальных водоемов" (хотя по крайней мере маты должны были существенную часть времени находиться в воде). Он считает, что "существование настоящих континентальных водоемов, как текучих, так и стоячих, представляется весьма проблематичным до того, как в девоне сосудистая растительность несколько снизила скорость эрозии и стабилизировала береговую линию". Основные события должны были происходить в уже знакомых нам выположенных прибрежных амфибиотических ландшафтах без стабильной береговой линии - "не суша, не море" (см. главу 5).
Не менее необычная (с точки зрения сегодняшнего дня) обстановка должна была складываться и на водоразделах, занятых "первичными пустынями". В наши дни пустыни существуют в условиях недостатка влаги (при превышении испарения над выпадением осадков) - что и препятствует развитию растительности. А вот в отсутствие растений ландшафт парадоксальным образом становился тем более пустынным (по облику), чем больше выпадало осадков: вода активно размывала горные склоны, прорезая глубокие каньоны, при выходе на равнину давала конгломераты, а дальше по равнине распространялись разрозненные по поверхности псефиты [33], которые называют равнинным пролювием; ныне такие отложения слагают лишь конусы выноса временных водотоков.
Эта картина позволяет по-новому взглянуть на одно странное обстоятельство. Почти все известные силурийско-девонские наземные флоры и фауны найдены в различных точках древнего Континента красного песчаника (Old Red Sandstone), названного так по характерным для него породам - красноцветам; все местонахождения при этом связаны с отложениями, считающимися дельтовыми. Иными словами, выходит, что весь этот континент (объединявший Европу и восток Северной Америки) представляет собой как бы одну сплошную гигантскую дельту. Резонный вопрос: а где же располагались соответствующие реки - ведь для них на континенте таких размеров просто нет площадей водосбора! Остается предположить что все эти "дельтовые" отложения, по всей видимости, возникали именно в результате процессов эрозии в описанных выше "мокрых пустынях".
Итак, жизнь на суше (которая, впрочем, не вполне еще суша) вроде бы существует с незапамятных времен, а в конце силура просто-напросто появляется еще одна группа растений - сосудистые (Tracheophyta)... Однако на самом деле появление сосудистых растений - одно из ключевых событий в истории биосферы, ибо по своей средообразующей роли эта группа живых организмов не имеет себе равных, по крайней мере, среди эукариот. Именно сосудистая растительность и внесла, как мы увидим далее, решающий вклад в становление наземных ландшафтов современного облика.
Общепринятой считается та точка зрения, что некие водоросли, жившие вблизи берега, сначала "высунули голову на воздух", затем заселили приливно-отливную зону, а потом, постепенно превращаясь в высшие растения, целиком вышли на берег. За этим последовало постепенное завоевание ими суши. Предками высших растений большинство ботаников считает одну из групп зеленых водорослей - харовых (Charophyta); они образуют ныне сплошные заросли на дне континентальных водоемов - как пресных, так и соленых, тогда как в море (да и то лишь в опресненных заливах) найдены всего несколько видов. Харовые обладают дифференцированным талломом и сложно устроенными органами размножения; с высшими растениями их сближают несколько уникальных анатомических и цитологических признаков - симметричные спермии, присутствие фрагмопласта (структуры, участвующей в построении клеточной стенки в процессе деления) и наличие одинакового набора фотосинтетических пигментов и запасных питательных веществ.
Однако против этой точки зрения выдвигалось серьезное - чисто палеонтологическое - возражение. Если процесс преобразования водорослей в высшие растения действительно происходил в прибрежных водах (где условия для попадания в палеонтологическую летопись наиболее благоприятны), то почему мы не видим никаких его промежуточных стадий? Тем более, что и сами харовые появляются в позднем силуре - одновременно с сосудистыми растениями, а особенности биологии этой группы не дают оснований оснований предполагать для нее длительный период "скрытого существования"... Поэтому появилась парадоксальная, на первый взгляд, гипотеза: а почему, собственно говоря, появление макроостатков высших растений в конце силура должно однозначно трактоваться как следы их выхода на сушу? Может быть, совсем наоборот - это следы переселения высших растений в воду? Во всяком случае, многие палеоботаники (С.В. Мейен, Г. Стеббинс, Г. Хилл) активно поддерживали гипотезу о происхождении высших растений не от водных макрофитов (типа харовых), а от наземных зеленых водорослей. Именно этим сухопутным (а потому не имеющим реальных шансов попасть в захоронения) "первичным высшим растениям" могли принадлежать загадочные споры с трехлучевой щелью, которые весьма многочисленны в раннем силуре и даже в позднем ордовике (начиная с карадокского века).
Впрочем, недавно выяснилось, что, судя по всему, правы сторонники обеих точек зрения - каждые по-своему. Дело в том, что некоторые из микроскопических наземных зеленых водорослей имеют тот же самый комплекс тонких цитологических признаков, что харовые и сосудистые (см. выше); эти микроводоросли теперь стали включать в состав Charophyta. Таким образом, возникает вполне логичная и непротиворечивая картина. Первоначально существовала - на суше - группа зеленых водорослей ("микроскопические харовые"), от которой в силуре произошли две близкородственные группы: "настоящие" харовые, заселившие континентальные водоемы, и высшие растения, начавшие осваивать сушу, и лишь спустя некоторое время (в полном соответствии со схемой Мейена) появившиеся в прибрежных местообитаниях.
Из курса ботаники вам должно быть известно, что высшие растения (Embryophyta) делятся на сосудистые (Tracheophyta) и мохообразные (Bryophyta) - мхи и печеночники. Многие ботаники (например, Дж. Ричардсон, 1992) считают, что именно печеночники (если исходить из их современных жизненных стратегий) - главные претенденты на роль "первопроходцев суши": они живут ныне на наземных водорослевых пленках, в мелководных эфемерных водоемах, в почве - совместно с синезелеными водорослями. Интересно, что азотфиксирующая сине-зеленая водоросль Nostoc способна жить внутри тканей некоторых печеночников и антоцеротовых, обеспечивая своих хозяев азотом; это наверняка было очень важно для первых обитателей примитивных почв, где этот элемент не мог не быть в жестком дефиците [34]. Упоминавшиеся выше споры из позднеордовикских и раннесилурийских отложений наиболее схожи именно со спорами печеночников (достоверные макроостатки этих растений появляются позже, в раннем девоне).
Однако в любом случае мохообразные (если даже они действительно появились еще в ордовике) облик континентальных ландшафтов вряд ли изменили. Первые же сосудистые растения - риниофиты [35] - появились в позднем силуре (лудловский век); вплоть до раннего девона (жединский век) они были представлены крайне однообразными остатками единственного рода Cooksonia, простейшего и архаичнейшего из сосудистых. А вот в отложениях следующего века девона (зигенского) мы находим уже множество разнообразных риниофитов (рисунок 30). С этого времени среди них обособляются две эволюционные линии. Одна из них пойдет от рода Zosterophylum к плауновидным (в их число входят и древовидные лепидодендроны - одни из основных углеобразователей в следующем, карбоновом, периоде). Вторая линия (в ее основание обычно помещают род Psilophyton) ведет к хвощеобразным, папоротникам и семенным - голосеменным и покрытосеменным (рисунок 30). Даже девонские риниофиты еще очень примитивны и, честно говоря, неясно - можно ли назвать их "высшими растениями" в строгом смысле: у них есть проводящий пучок (правда, сложенный не трахеидами, а особыми вытянутыми клетками со своеобразным рельефом стенок), но отсутствуют устьица. Такое сочетание признаков должно свидетельствовать о том, что растения эти никогда не сталкивались с дефицитом воды (можно сказать, что вся их поверхность являет собою одно большое открытое устьице), и, по всей видимости, являлись гелофитами (то есть росли "по колено в воде", вроде нынешнего камыша).
 |
| РИСУНОК 30. Родословное древо первых наземных растений. |
Появление сосудистых растений с их жесткими вертикальными осями вызвало целый каскад экосистемных новаций, изменивших облик всей биосферы:
1. Фотосинтезирующие структуры стали располагаться в трехмерном пространстве, а не на плоскости (как это было до сих пор - в период господства водорослевых корок и лишайников). Это резко увеличило интенсивность образования органического вещества и, тем самым, суммарную продуктивность биосферы.
2. Вертикальное расположение стволов сделало растения более устойчивыми к занесению смываемым мелкоземом (по сравнению, например, с водорослевыми корками). Это уменьшило безвозвратные потери экосистемой неокисленного углерода (в виде органики) - совершенствование углеродного цикла.
3. Вертикальные стволы наземных растений должны быть достаточно жесткими (по сравнению с водными макрофитами). Для обеспечения этой жесткости возникла новая ткань - древесина, которая после гибели растения разлагается относительно медленно. Таким образом, углеродный цикл экосистемы обретает дополнительное резервное депо и, соответственно, стабилизируется.
4. Появление постоянно существующего запаса трудноразложимой органики (сконцентрированного в основном в почве) ведет к радикальной перестройке пищевых цепей. С этого времени большая часть вещества и энергии оборачивается через детритные, а не через пастбищные цепи (как это было в водных экосистемах).
5. Для разложения трудноусваиваемых веществ, из которых состоит древесина - целлюлозы и лигнина - потребовались новые типы разрушителей мертвой органики. С этого времени на суше роль основных деструкторов переходит от бактерий к грибам.
6. Для поддержания ствола в вертикальном положении (в условиях действия силы тяжести и ветров) возникла развитая корневая система: ризоидами - как у водорослей и мохообразных - тут уже не обойдешься. Это привело к заметному снижению эрозии и появлению закрепленных (ризосферных) почв.
С.В. Мейен полагает, что суша должна была покрыться растительностью к концу девона (зигенский век) [36], поскольку с начала следующего, карбонового, периода на Земле образуются уже практически все типы осадков, отлагающихся ныне на континентах. В дозигенские же времена континентальные осадки практически отсутствуют - видимо, в связи с их постоянным вторичным размывом в результате нерегулируемого стока. В самом начале карбона на континентах начинается угленакопление - а это свидетельствует о том, что на пути стока вод стояли мощные растительные фильтры. Не будь их, остатки растений непрерывно смешивались бы с песком и глиной, так что получались бы обломочные породы, обогащенные растительными остатками - углистые сланцы и углистые песчаники, а не настоящие угли.
Итак, возникшая в прибрежных амфибиотических ландшафтах густая "щетка" из гелофитов (можно назвать ее "риниофитный камыш") начинает действовать как фильтр, регулирующий плащевой сток: она интенсивно отцеживает (и осаживает) сносимый с суши обломочный материал и формирует тем самым стабильную береговую линию. Некоторым аналогом этого процесса может служить формирование крокодилами "аллигаторовых прудов": животные постоянно углубляют и расширяют населяемые ими болотные водоемы, выбрасывая грунт на берег. В результате их многолетней "ирригационной деятельности" болото превращается в систему чистых глубоких прудов, разделенных широкими облесенными "дамбами". Так и сосудистая растительность в девоне разделила пресловутые амфибиотические ландшафты на "настоящую сушу" и "настоящие пресноводные водоемы". Не будет ошибкой сказать, что именно сосудистая растительность стала истинным исполнителем заклинания: "Да будет твердь!" - отделивши оную твердь от хляби...
Именно с вновь возникшими пресноводными водоемами связано и появление в позднем девоне (фаменский век) первых тетрапод (четвероногих) - группы позвоночных, имеющих две пары конечностей; она объединяет в своем составе амфибий, рептилий, млекопитающих и птиц (попросту говоря, тетраподы - это все позвоночные, кроме рыб и рыбообразных). В настоящее время общепринято, что тетраподы ведут свое происхождение от кистеперых рыб (Rhipidistia) (рисунок 31); эта реликтовая группа имеет ныне единственного живого представителя, латимерию [37]. Достаточно популярная некогда гипотеза происхождения четвероногих от другой реликтовой группы рыб - двоякодышащих (Dipnoi), ныне практически не имеет сторонников.
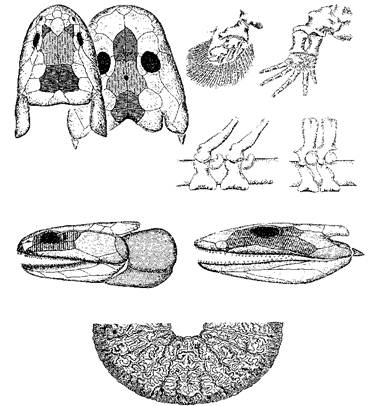
РИСУНОК 31. Преемственность анатомических структур (череп, конечности и позвонки) кистеперых рыб и амфибий: идентичные кости черепа имеют одинаковую заливку; идентичные кости обозначены одинаковыми буквами; отдельно - поперечный шлиф половинки зуба лабиритнтодонта.
Необходимо отметить, что в прежние годы возникновение ключевого признака тетрапод - двух пар пятипалых конечностей - считали их однозначной адаптацией к наземному (или по меньшей мере амфибиотическому) образу жизни. Ныне, однако, большинство исследователей склоняется к тому, что "проблема появления четвероногих" и "проблема их выхода на сушу" - суть вещи разные и даже не связанные между собою прямой причинной связью. Предки тетрапод жили в мелководных, часто пересыхающих, обильно заросших растительностью водоемах непостоянной конфигурации. Судя по всему, конечности появились для того, чтобы передвигаться по дну водоемов (это особенно важно, когда водоем обмелел настолько, что у тебя уже спина начинает вытарчивать наружу) и продираться сквозь плотные заросли гелофитов; особенно же полезны конечности оказались для того, чтобы при пересыхании водоема переползти посуху в другой, соседний.
Первые, девонские, тетраподы - примитивные амфибии лабиринтодонты [38] (название это происходит от их зубов с лабиринтоподобными складками эмали - структура, прямо унаследованная от кистеперых: см. рисунок 31), такие как ихтиостега и акантостега, в захоронениях всегда встречаются вместе с рыбами, которыми, судя по всему, и питались. Они были покрыты чешуей, как рыбы, имели хвостовой плавник (похожий на тот, что мы видим у сома или налима), органы боковой линии и - в некоторых случаях - развитый жаберный аппарат; конечность их еще не пятипалая (число пальцев достигает 8), и по типу сочленения с осевым скелетом - типично плавательная, а не опорная. Все это не оставляет сомнения в том, что существа эти были чисто водными (рисунок 32); если они и появлялись на суше при неких "пожарных" обстоятельствах (пересыхание водоема), то компонентом наземных экосистем наверняка не являлись [39]. Лишь много позже, в карбоновом периоде, появились мелкие наземные амфибии - антракозавры, которые, судя по всему, питались членистоногими, но об этом речь впереди (см. главу 10).

РИСУНОК 32. Архаичный лабиринтодонт Acanthostega: скелет (а) и реконструкция в естественной обстановке (б).
Особого внимания заслуживает тот факт, что в девоне появляется целый ряд неродственных параллельных групп стегоцефалоподобных кистеперых рыб - причем как до, так и после возникновения "настоящих" тетрапод (лабиринтодонтов). Одной из таких групп были пандерихтиды - кистеперые, лишенные спинного и анального плавников, чего не бывает ни у каких других рыб. По строению черепа (уже не "рыбьему", а "крокодильему"), плечевого пояса, гистологии зубов и положению хоан (внутренних ноздрей) пандерихтиды очень сходны с ихтиостегой, однако приобрели эти признаки явно независимо. Таким образом, перед нами процесс, который можно назвать параллельной тетраподизацией кистеперых (он был детально изучен Э.И. Воробьевой). Как обычно, "заказ" на создание четвероногого позвоночного, способного жить (или, по крайней мере, выживать) на суше был дан биосферой не одному, а нескольким "конструкторским бюро"; "выиграла конкурс" в конечном итоге та группа кистеперых, которая "создала" известных нам тетрапод современного типа. Однако наряду с "настоящими" тетраподами еще долго существовал целый спектр экологически сходных полуводных животных (типа пандерихтид), сочетающих признаки рыб и амфибий - если так можно выразиться, "отходы" процесса тетраподизации кистеперых.
Выдающийся палеоботаник С.В. Мейен, анализируя механизм возникновения прочно устоявшихся научных предрассудков, приводит в качестве примера цитату из школьного учебника биологии, где речь идет о карбоновом (каменноугольном) периоде "с его теплым влажным климатом и воздухом, богатым углекислым газом вследствие сильной вулканической деятельности... С конца каменноугольного периода в связи с усиленным горообразованием, охватившим в следующем (пермском) периоде весь земной шар, влажный климат почти повсеместно сменился сухим. В новых условиях древовидные папоротники стали быстро вымирать... Вымерли и семенные папоротники." Дело даже не в прямых фактических ошибках (например, "семенные папоротники" [40], которые якобы погибли из-за установления в перми засушливого климата, на самом деле благополучно дожили почти до конца мезозоя, а пермь вообще была временем их расцвета). Серьезнее другое: представления о том, что карбон - время теплого и влажного климата, а пермь - время повсеместной аридизации, сформировались в прошлом веке; эта картина была нарисована европейскими геологами и палеонтологами на европейском материале, а затем - безо всяких на то оснований! - распространена на всю Землю. Уже в начале нашего века стало ясно, что ситуация в Еврамерийской области карбона (территория нынешних Европы и Северной Америки) сходна с Катазией (Китай и Индокитай), но радикально отлична от того, что наблюдалось в Гондване (на материках Южного полушария и Индии) и в Ангариде (северная Азия); тем не менее, на возникший чуть ли не полтора века назад стереотип все это ничуть не повлияло - как можно видеть из цитированного выше учебника 80-х годов.
Причины климатических различий, существовавших в позднем палеозое между Еврамерией и Катазией, с одной стороны, и Гондваной и Ангаридой - с другой, кажутся вполне очевидными: первые располагались на тогдашнем экваторе, а вторые - вблизи полюсов. Представьте-ка себе, что мы попытаемся экстраполировать на всю нынешнюю Землю картину амазонских джунглей!.. Однако здесь сразу же возникает встречный вопрос: а всегда ли на Земле существовала широтная климатическая зональность, сходная с нынешней? Для ответа логично обратиться к сопоставлению высоко- и низкоширотных флор соответствующих эпох прошлого (учитывая при этом иное, чем теперь, расположение материков относительно полюса).
Картина эволюции растительности от девона до наших дней выяснена (в общих чертах) достаточно давно. Для наиболее молодых, кайнозойских, флор установлена ясная климатическая зональность, хотя и отличная от нынешней (на арктических островах росли деревья, характерные ныне для зоны широколиственных лесов - например, каштаны и платаны). Мезозойские флоры существенно более однообразны по всей Земле. Сложнее ситуация с палеозоем. Пермские и позднекарбоновые флоры Европы и Северной Америки, как уже было сказано, сходны с китайскими, но резко отличны и от сибирских, и от флор всех материков Южного полушария. Но ниже по геологическому разрезу - в раннем карбоне и далее, в девоне, мы снова сталкиваемся с единством флор различных материков. Отсюда можно заключить, что климатическая зональность была минимальной в девоне и начале карбона, затем усилилась в позднем палеозое, снова ослабла в мезозое, а потом опять начала усиливаться, достигнув ныне своего максимума.
Мы с вами помним, что в верхнепалеозойских слоях всех гондванских материков найдены ледниковые отложения - тиллиты (которые послужили одним из отправных пунктов в построениях Вегенера); следы оледенений найдены и в одновозрастных отложениях Ангариды. А вот за изъятием этого отрезка времени (поздний карбон - ранняя пермь) и современности климат в высоких широтах был если и не жарким, то во всяком случае безморозным: в раннем карбоне и Европы, и Шпицбергена, и Сибири найдены толстые, явно многолетние стволы плауновидных с маноксилической древесиной [41], а в эоцене острова Элсмир (Канадский архипелаг) - крокодилы. Современная климатическая картина с крупными полярными шапками из снега и льда - скорее исключение, чем правило в геологической истории. Так что следует искать ответа не на вопрос, почему в раннем карбоне и в мезозое не было полярных шапок, а на вопрос, отчего они иногда образовывались (и меняли весь климат планеты). Периоды существования такого контрастного климата с холодными полюсами, как в позднем палеозое и позднем кайнозое, называют криоэрами ("криос" - по-гречески холод), а выровненного по всей Земле (как в мезозое) - соответственно, термоэрами. Общее количество тепла, получаемое Землей от Солнца, считается достаточно постоянным во все времена; здесь существует своя циклика (см. главу 14), но расстояние-то между ними неизменно. Следовательно, дело в основном в распределении этого тепла по поверхности планеты, прежде всего - в характере и интенсивности теплопереноса от экватора к полюсам.
Для начала, как водится, несколько общих замечаний. Поскольку планета шарообразна, солнечные лучи всегда будут, при прочих равных, нагревать ее экват








