2013-12-31
2013-12-31 1071
1071ПОБЕГ
Классификация по Раункиеру
Зоогенные факторы
Это воздействие животных друг на друга и окружающую среду. К ним относится также потребление животными растительной пищи. Такие животные называются фитофагами (от греч. phyton — растение и phagos — пожирающий). Фитофагами являются крупные (лось, олень, косуля, кабан) и мелкие (заяц, белка, мышевидные грызуны) млекопитающие, птицы (рябчик, тетерев, глухарь), насекомые-вредители и др.
Контактируя с растениями или употребляя их в пищу, животные способствуют распространению их семян. В одних случаях семена и плоды распространяются благодаря случайному прикреплению к животным (шерсти, перьям, лапам, клювам и т.п.), в других это связано с поеданием животными плодов. Животные серьезно повреждают растения. Лоси и олени обдирают кору на деревьях, уничтожают молодую древесную поросль, объедая верхушки кустарников и древесного подроста. Бобры, питаясь древесиной осины, довольно быстро изреживают ее насаждения. Глухари, ощипывая хвою и почки сосны и ели, тем самым замедляют их рост.
К зоогенным факторам относится воздействие насекомых на листовую поверхность древесных пород и травянистых растений. Насекомые (тли, клопы) не только отсасывают у растений питательные вещества, но и переносят возбудителей их заболеваний.
Большой вред растениям наносят землерои (кроты, суслики). Они поедают не только надземные части растений, но и клубни, луковицы, корневища.
Воздействие животных на растения довольно многообразно и сказывается на регулировании численности видов в природных сообществах.
Действие зоогенных факторов в среде животных проявляется главным образом в виде паразитизма, хищничества и конкуренции.
экологическая классификация растений, основанная на положении почек возобновления и наличии соответствующих приспособлений для переживания неблагоприятного времени года. По системе раункиера различают 6 групп жизненных форм: эпифиты — растения, не имеющие корней в почве; фанерофиты — надземные растения, почки возобновления которых находятся на вертикально расположенных побегах; хамефиты — поверхностные растения, почки возобновления которых расположены высоко над землей, гемикриптофиты — дернообразующие растения, почки возобновления которых находится на уровне почвы или непосредственно под ее поверхностью; криптофиты — растения, зимующие клетки, почки возобновления которых скрыты в почве (геофиты) или под водой (гидрофиты); терофиты — однолетние растения, жизненный цикл которых заканчивается за один вегетационный сезон (неблагоприятное время года переживают в виде семян)
ЭПИФИТЫ
[от эпи... и...фит(ы)], живущие на другие растениях (главным образом на стволах и ветвях) непаразитарные “воздушные растения”, не имеющие корней в почве. Поглощают атмосферную влагу через все тело или через воздушные корни. Эпифиты не имеют прямого физиологического или биохимического контакта с хозяином, поскольку живут не за счет его самого, а используя мелкозем и воду, скопившуюся в неровностях на стволах и ветвях. Лишь отчасти эпифиты прибегают к продуктам распада самых верхних частей коры. Примеры эпифитов — орхидеи, мхи, лишайники и др.
ФАНЕРОФИТЫ
[от греч. phaneros — явный, открытый и...фит (ы)] растения, почки и концевые побеги которых находятся высоко над поверхностью почвы и переживают неблагоприятное время года без особой защиты. К фанерофиты относятся деревья и кустарники, стеблевые суккуленты, лианы и др.
ГЕМИКРИПТОФИТЫ
(от геми... и криптофиты), растения, у которых почки возобновления в неблагоприятный для вегетации период года сохраняются на уровне почвы (иногда чуть выше) и защищены чешуями, опавшими листьями и снежным покровом. К гемикриптофитам относятся очень многие травянистые растения средних широт (лютик, живучка ползучая, одуванчик и др.)
Хамефиты
[от греч. chamai — на земле и...фит(ы)], “поверхностные растения”, мелкие кустарники, полукустарники (брусника, черника, вереск) и травянистые растения (стелющиеся формы), почки возобновления которых находятся на слабо возвышающихся над почвой побегах. Почки защищены от действия неблагоприятных факторов почечными чешуями, снегом (зимой) и подстилкой. Хамефиты чаще всего встречаются в тундрах, высокогорьях, пустынях
КРИПТОФИТЫ
[от греч. kryptos — тайный, скрытый и...фит(ы)], жизненная форма растений, у которых почки возобновления закладываются на корневищах, клубнях, луковицах и находятся в почве (геофиты) или под водой (гидрофиты)
ТЕРОФИТЫ
[от греч. theros — лето и...фит(ы)], жизненная форма в системе Раункиера, означающая однолетние растения, полностью отмирающие к зиме, но сохраняющие жизнеспособные семена. Это однолетние травы средиземноморского происхождения, характерные для пустынь, полупустынь, степей (маковые, крестоцветные и др.)
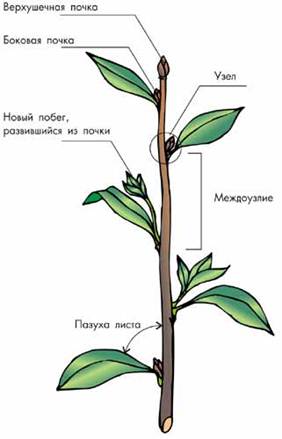
Побег – основной орган высших растений.
Основные элементы побега:
1) Ось побега – стебель. Самые длинные стебли, длиной свыше 100 м, характерны для древесных растений: секвойи, мамонтова дерева (секвойядендрона), произрастающих в Северной Америке. Среди лиан наиболее длинные стебли у ротанговой пальмы (до 400 м длиной). Самые низкорослые стебли (около 2 мм) – у австралийских орхидей.
Толщина стеблей (стволов) может достигать значительных размеров. Например, у секвойи - до 36 м в диаметре, толщина корки – 70-80 см. На пне одной срубленной секвойи была устроена танцплощадка, внутри другого поваленного дерева построили ресторан на 50 мест, еще одно приспособили под гараж.
Долгожителями называют: мексиканский кипарис (живет 10 тыс. лет), баобаб (до 6 тыс. лет).
Функции стебля:
· место образования листьев и цветков, а в их пазухах (угол между стеблем и листом) - пазушных почек;
·  ассимиляция органических веществ (у некоторых растений только стебли выполняют функцию фотосинтеза (например, у кактусов, хвощей);
ассимиляция органических веществ (у некоторых растений только стебли выполняют функцию фотосинтеза (например, у кактусов, хвощей);
· транспорт воды, минеральных и органических веществ из корня к листьям и обратно;
· запасание питательных веществ и воды; вегетативное размножение;
· защита растений (развиваются колючки и шипы).
2) Стебель состоит из узлов и междоузлий:
· участок стебля, от которого отходит лист (или листья), принято называть узлом.
·  участки стебля между соседними узлами — междоузлия. Каждый повторяющийся элемент побега (междоузлие и узел с листом и почкой) называется метамером (рис. 1).
участки стебля между соседними узлами — междоузлия. Каждый повторяющийся элемент побега (междоузлие и узел с листом и почкой) называется метамером (рис. 1).
3) Первый побег растения — это главный побег, или побег первого порядка, образуется из зародышевого стебелька. Из боковых почек главного побега формируются побеги второго порядка, а при повторении ветвления — третьего и т. д.
4) Различают следующие типы побегов: вегетативные (состоят из стебля и листьев), спороносные - несущие репродуктивные органы (у покрытосеменных – цветки, у голосеменных – шишки), зачаточные - почки (содержат в зачаточном состоянии стебель, листья и генеративные органы).
5) Почка — это зачаточный побег.
Различают следующие типы почек:
I. По содержанию:
· вегетативные – содержат в зачаточном состоянии вегетативные органы – стебель и листья;
· генеративные - содержат в зачаточном состоянии цветки или соцветия. Цветочная почка, из которой развивается отдельный цветок, получила название бутона;
· вегетативно-генеративные - помимо вегетативных органов, заложены соцветие или отдельный цветок (например, у сирени).
Внутри почки заключена меристематическая верхушка побега — апикальная меристема (апекс, конус нарастания).
Почки, особенно у древесных растений зон холодного и умеренного климата, обычно окружены специализированными почечными чешуями, представляющими видоизмененные листья.
II. По наличию или отсутствию почечных чешуй:
· закрытые почки - прикрыты чешуями, выполняющими защитную функцию у древесных и кустарниковых растений зон умеренного и холодного климата.Наружные почечные чешуи растут слабо, постепенно засыхают и опадают, оставляя почечные кольца (рис. 1).
· открытые почки - у тропических растений, а также у травянистых почечные чешуи отсутствуют.
III. По расположению на побеге различают:
· верхушечные почки, осуществляют его рост.
· боковые, или пазушные, почки дают побеги следующего порядка. Они располагаются всегда в пазухе кроющего листа. На месте кроющего листа после его опадения на стебле образуется различной формы листовой рубец;
· спящие почки, расположеныу оснований стволов деревьев. В естественных условиях из спящих почек образуется корневая и пнёвая поросль — после отмирания или рубки леса;
· придаточные, или адвентивные почки, в отличие от спящих, могут формироваться на всех частях и органах растения, включая и корни.
ОТЛИЧИЯ В СТРОЕНИИ СТЕБЛЯ И КОРНЯ
Стебель и корень закладываются в зародыше как единая структура. Однако они отличаются друг от друга по строению и функциям:
1. первичная кора стебля покрыта эпидермой с устьицами, а в корне – ризодермой с корневыми волосками;
2. под эпидермой стебля расположена колленхима, а в корне ее нет;
3. в составе первичной коры стебля находятся хлоропласты (первичная кора выполняет функцию хлоренхимы), в корне их нет. Хлоренхима стебля образует чередующиеся продольные полосы с колленхимой и или склеренхимой (стебель укропа);
4. проводящая система более сложно устроена в стебле:
а) у некоторых хвойных и двудольных растений первичные проводящие ткани образуют сплошной цилиндр с воздушной полостью в центре;
б) у других хвойных и двудольных первичные проводящие ткани образуют цилиндр из отдельных проводящих пучков;
в) у большинства однодольных и некоторых травянистых двудольных проводящие пучки расположены хаотически.
5. В стеблях - проводящие пучки коллатерального типа (флоэма залегает над ксилемой (см. конспект «Проводящие ткани»), а в корне – радиального типа.
Анатомические особенности стеблей водных растений
Водная среда обитания растений характеризуется следующими факторами:
· ослабленное освещение,
· бедность среды диоксидом углерода и кислородом,
· отсутствие иссушающих факторов из-за постоянного обеспечения растений водой,
· механическая поддержка со стороны воды и др.
Эти факторы влияют на анатомическое строение водных растений. Особенности анатомического строения стеблей водных растенийследующие:
1. кожица слабо дифференцирована;
2. часто клетки кожицы содержат хлорофилловые зерна и способны к фотосинтезу;
3. отсутствует устьичный аппарат или он есть, но слабо развит;
4. первичная кора занимает большую часть стебля;
5. первичная кора построена из рыхлой тонкостенной паренхимы, с широкими воздухоносными ходами, располагающимися в один или несколько кругов и отделяющимися один от другого тонкими однослойными перегородками клеток; 6) небольшой по размеру центральный цилиндр;
6. сердцевина слабо выражена или отсутствует;
7. сосуды вскоре после образования могут разрушиться и в центре стебля остается воздухоносный ход;
8. слабо одревесневают лишь стенки сосудов;
9. крупные межклетники заполнены воздухом, сильно развита аэренхима.
ВЕТВЛЕНИЕ
Ветвление – генетически обусловленный признак, связанный с расположением боковых почек в апикальной меристеме побега. Если почки не закладываются или не развиваются, растения не ветвятся (пальмы, агавы, алоэ и др.). В этом случае растения имеют крупные листья.
материнских и дочерних осей (рис. 2):
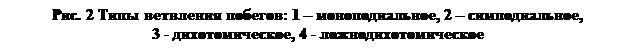
1. Моноподиальное ветвление (моно - один, подиум – ось) – при этом типе ветвления верхушечная почка в течение всей жизни растения продолжает рост главного стебля, который удлиняется и утолщается в большей степени, чем боковые ветви (у ели, сосны и др.).
2. Симподиальное ветвление (в переводе – составная ось) – при этом типе ветвления верхушечная почка со 2-5 –го года жизни растения замирает, а ближайшая к ней боковая почка дает побег, продолжающий рост стебля. Т.е. происходит перевершинивание, далее оно происходит каждый год. Чаще встречается у древесных и кустарниковых растений (береза, яблоня, грецкий орех и др.).
3. Дихотомическое ветвление характерно для мхов, плаунов, водорослей. Верхушечная инициальная (самая первая) клетка конуса нарастания побега или другого осевого органа дает начало двум осям, которые продолжают ветвиться вильчато.
4. Ложнодихотомическое ветвление характерно для цветковых растений, когда 2 ветви формируются из двух супротивно расположенных почек, а верхушечная почка замирает (сирень, конский каштан и др.).
ЛИСТОРАСПОЛОЖЕНИЕ, ЛИСТОВАЯ МОЗАИКА
Листорасположение - это порядок размещения листьев на оси побега. Листорасположение всегда симметрично и подчиняется правилу эквидистантности – равенству угловых расстояний между листьями, сидящими на одном узле или на последовательных узлах основной генетической спирали (условной спиральной линии, которой можно соединить основания последовательных листьев).
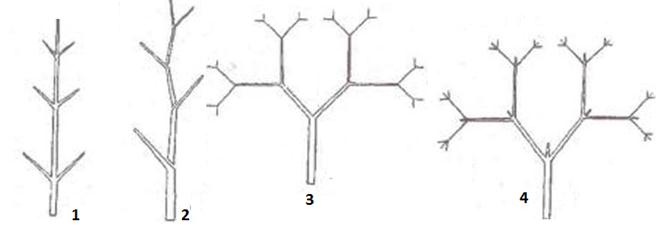
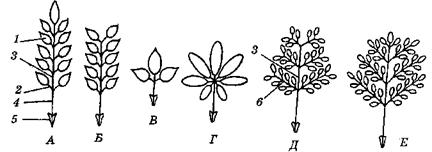
Основные типы листорасположения (рис. 3):
· спиральное, или очередное, — когда от каждого узла стебля отходит один лист (дуб, береза, злаки),
· частный случай очередного листорасположения – двурядное (узел закрытый, апикальная меристема откладывала листовые зачатки по принципу маятника, узлы сближены, все листья лежат в одной плоскости (у гладиолуса)),
· мутовчатое — когда каждый узел несет три и более листьев (элодея),
· супротивное — когда на каждом узле сидят друг против друга два листа (сирень).

В процессе роста побега черешки листьев могут изгибаться, и в силу этого положение пластинок изменяется. При этом пластинки всех листьев в конечном итоге располагаются, не затеняя друг друга, но образуют единую плоскость, где просветы между крупными листьями заполнены более мелкими листьями. Подобное явление, получившее название листовой мозаики, позволяет растению более полно использовать падающий на него солнечный свет.
ЛИСТ
Лист, в отличие от корня и стебля, - боковой орган ограниченного роста. Нарастают листья не верхушкой, а при основании, т.е. интеркалярно. Исключение составляют листья папоротников – вайи, удлинение которых идет за счет действия верхушечной (апикальной) меристемы, что говорит об их стеблевом происхождении.
У большинства растений лист – это временный орган (исключение – южноафриканское растение вельвичия, у которой 2 супротивных листа отмирают в конце жизни самого растения в возрасте 90-100 лет). Среднее время жизни листьев 45-120 дней. От 2 до 5 лет живут листья у ряда так называемых вечнозеленых растений субтропиков и тропиков, а также у некоторых растений тайги. У части хвойных продолжительность жизни листа достигает 15-20 лет.
Размеры листьев чаще всего колеблются в пределах от 3 до 15 см, однако известны гигантские листья некоторых пальм рода Рафия - до 22 м в длину и 12 м в ширину. Из водных растений самые крупные листья (2 м в диаметре) имеет виктория королевская. Размеры, форма и степень рассеченности листьев, хотя и являются наследственными признаками того или иного вида, очень изменчивы и зависят также от условий обитания растения.
Функции листа:
1. фотосинтез;
2. газообмен и транспирация;
3. синтез органических веществ – фитогормонов (гиббереллинов, абсцизовой кислоты и др.);
4. запасающая функция (например, запасание воды в мясистых сочных листьях алоэ и др. растений);
5. листья, видоизмененные в иголки, защищают растение от перепадов температуры, избыточного испарения и фитофагов;
6. листья, видоизмененные в усики, способствуют прикреплению растения к опоре;
7. листья, видоизмененные в ловчий аппарат, служат для захвата насекомых, иногда мелких земноводных и др. животных. Они содержат парализующее вещество (например, кониин), а также муравьиную кислоту, подавляющую рост гнилостных бактерий.
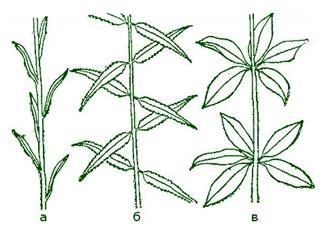
Морфология листа
Взрослый лист (рис.4) обычно расчленен на пластинку или несколько пластинок (у сложных листьев) и черешок — узкую стеблевидную его часть, соединяющую пластинку и узел побега. Самая нижняя часть листа, сочлененная со стеблем, называется основанием листа. Часто при основании заметны разного размера и формы парные боковые выросты — прилистники.
У двудольных растений лист, как правило, — плоский орган, снабженный черешком. Кроме опорной и проводящей функций, он может регулировать положение пластинки, изгибаясь по направлению к свету (формирует листовую мозаику). Нередко черешок не развивается, и тогда лист называют сидячим. Лист с черешком называют черешковым.
| 
|
Основание листа принимает различную форму. Однако нередко, особенно у ряда однодольных (например, у злаков и зонтичных), оно разрастается и образует замкнутую или незамкнутую трубку, называемую листовым влагалищем. Листовое влагалище защищает пазушные почки и нередко служит средством дополнительной опоры побега.
Простые и сложные листья (рис. 5)
1. Лист, имеющий одну пластинку, называется простым.
2. У сложного листа две, три или большее число обособленных пластинок (листочков), снабженных собственными черешочками и иногда особыми сочленениями. Часть оси сложного листа, несущую листочки, называют рахисом (от греч. позвоночник). В зависимости от расположения листочков различают:
а) перистосложные листья - листочки располагаются двумя рядами по обе стороны рахиса, продолжающего черешок. По степени разветвления рахиса различают однократно, дважды и трижды перистосложныелистья. Если рахис любого порядка перистосложного листа завершается на верхушке непарным листочком, лист является непарно-перистосложным, а при отсутствии верхушечного листочка — парно-перистосложным. Трижды непарно-перистосложный тип листа известен лишь у одного растения — тропического вида моринги.. Дважды парно-перистосложные листья весьма обычны у представителей подсемейства мимозовых (сем. Бобовые). Число мелких листочков такого листа иногда достигает 10 тысяч.
б) пальчатосложные листья и их частный случай — тройчатосложные листья не имеют рахиса, и листочки отходят от верхушки черешка. Классический пальчатосложный лист у видов конского каштана винограда девичьего пятилисточкового, в просторечии называемого диким виноградом. Представление о тройчатосложном листе можно составить, вспомнив листья клевера, земляники.

Жилкование листьев (рис. 6)
В листе проходят проводящие пучки, которые обычно различным образом ветвятся и образуют анастомозы – перемычки между крупными жилками. В большинстве случаев проводящая система листа выступает в виде так называемого жилкования, обычно особенно хорошо выраженного на нижней его стороне.
Проводящая система листа цветковых растений обычно представляет собой довольно сложную картину, в которой более или менее ясно выделяются жилки разного порядка ветвления. Жилки первого порядка представляют собой самые толстые, главные жилки листа. От них отходят более тонкие жилки второго и последующих порядков.
Типы жилкования листьев:
1. Перистое жилкование представлено одной главной жилкой, от которой отходят боковые жилки второго порядка. Этот тип жилкования характерен для двудольных растений. Различают:
а) перистокраевое жилкование, когда боковые жилки тянутся прямо до края пластинки и оканчиваются здесь в лопастях, концах зубчиков, выемках или даже выступают в виде щетинок. Оно встречается, например у березы, вяза.
б) перистопетлевое жилкование, когда боковые жилки направляются к краю пластинки, но, еще не достигнув его, образуют петлю и возвращаются к главной жилке. Такое жилкование характерно для довольно большого числа двудольных, в том числе для лавра (лавровый лист), молочая и других растений.
в) перистосетчатое жилкование, когда жилки второго порядка не доходя по края листа, многократно ветвятся, образуя густую сеть, ясно выраженные петли здесь отсутствуют. Такое жилкование хорошо выражено у видов ивы, груши, яблони, айвы.


2.Пальчатое жилкование - другая разновидность жилкования листьев у двудольных растений. Первичные жилки отходят радиально от одной точки или у самого основания пластинки или несколько выше основания. Примером такого жилкования могут служить листья клена. В пальчатом жилковании повторяются те же основные типы, которые мы видели у перистого жилкования:
а) пальчатокраевое жилкование (например, у клена, винограда),
б) пальчатопетлевое жилкование (например, у церцидифиллума),
в) пальчатосетчатое (например, лукосемянника).
3.Дуговидное жилкование. Главные жилки вступают по нескольку в пластинку, причем внешние из них направляются параллельно краю пластинки дугой к вершине листа. Жилки второго порядка, образуют перемычки, соединяющие соседние главные жилки. Подобное жилкование встречается у многих однодольных, например, у ландыша подорожника.
4. Параллельное жилкование. Первичные жилки (две или больше) входят самостоятельно из листового влагалища в основание пластинки и идут далее более или менее параллельно до верхушки листа, где смыкаются. Параллельные первичные жилки соединены тонкими прямыми или косыми перемычками. Типичное параллельное жилкование можно видеть у многих лилейных, осок и особенно злаков.
5. Дихотомическое (вильчатое) жилкование – редкий тип жилкования. Распространено у папоротников и архаичных (от греч. архос – древний) растений (например, гинкго).
АНАТОМИЧЕСКОЕ СТРОЕНИЕ ЛИСТА
Строение листа двудольного растения на примере камелии
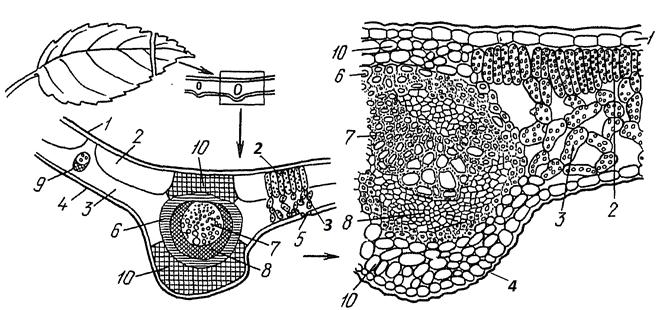 На срезе листа сверху и снизу расположена покровная ткань — эпидерма (Рис. 7). Основными отличительными признаками верхней эпидермы по сравнению с нижней, являются: более утолщенная наружная клеточная стенка, более мощная кутикула на поверхности и почти полное отсутствие устьиц на верхней эпидерме.
На срезе листа сверху и снизу расположена покровная ткань — эпидерма (Рис. 7). Основными отличительными признаками верхней эпидермы по сравнению с нижней, являются: более утолщенная наружная клеточная стенка, более мощная кутикула на поверхности и почти полное отсутствие устьиц на верхней эпидерме.
 Между верхней и нижней эпидермой находится ткань, в клетках которой содержится хлорофилл. Это ассимиляционная ткань листа — мезофилл.
Между верхней и нижней эпидермой находится ткань, в клетках которой содержится хлорофилл. Это ассимиляционная ткань листа — мезофилл.
Клетки мезофилла, примыкающие к верхней эпидерме, имеют вытянутую форму, плотно сомкнуты, без межклетников, расположены в два слоя. Это – столбчатый мезофилл или столбчатая (палисадная) паренхима. Здесь в основном происходит фотосинтез.
У нижней эпидермы расположены более округлые клетки с крупными межклетниками — это губчатый мезофилл (или губчатая паренхима). Главная функция нижней стороны листа — газообмен и транспирация.
В некоторых клетках губчатой паренхимы могут находиться друзы оксалата кальция, а также крупные разветвленные клетки — склереиды, выполняющие опорную функцию.
Между клетками мезофилла на некотором расстоянии друг от друга 999 тонкие жилки листа, содержащие проводящие ткани (ксилему и флоэму). Главная жилка занимает почти всю толщу листа - от верхней до нижней эпидермы. При малом увеличении хорошо видна мощная ксилема. К ксилеме непосредственно примыкает флоэма. Особенностью их расположение является то, что ксилема всегда обращена к верхней стороне листа, а флоэма — к нижней. Пучок окружен слоем склеренхимы.
АНАТОМО-МОРФОЛОГИЧЕСКИЕ ОСОБЕННОСТИ ЛИСТЬЕВ ВОДНЫХ РАСТЕНИЙ
1. Листья погруженных гидрофитов очень тонки и нежны, имеют упрощенное строение мезофилла без заметной дифференциации на палисадную и губчатую паренхиму.
2. Подводные листья без устьиц.
3. В отдельных местах находятся группы клеток эпидермиса с утонченными стенками. Считается, что они играют большую роль в поглощении воды и растворенных минеральных солей.
4. У растений лишь частично погруженных в воду, хорошо выражена гетерофиллия - различие строения надводных и подводных листьев на одном и том жерастений. Первые имеют черты, обычные для листьев наземных растений, вторые – очень тонкие или рассеченные листовые пластинки. Гетерофиллия отмечена у водного лютика – Ranunculus diversifolius, кувшинок и кубышек.