2020-04-20
2020-04-20 239
239
В момент детерминации клетки специализирующиеся на выполнении функций пигментообразования получают название меланобластов. Они делятся и начинают мигрировать из нервной трубки к местам назначения в эпидермис, туда, где их поджидают дифференцирующиеся волосяные фолликулы. Однако этот процесс может быть прерван в самом начале, если хотя бы одна из хромосом несет доминантную мутацию W. Животные, гомо - (W/W) и гетерозиготные (W/+) по этой мутации, имеют белую окраску, так как меланобласты не могут дойти до волос.
Эффект гена W может служить хорошим примером явления эпистаза, или подавления эффектов неаллельных генов: генетически рыжие и генетически серые коты, имея в своем генотипе мутацию W, будут одинаково белыми. Еще одно генетическое явление иллюстрирует собой ген W-плейотропию, или множественное действие гена. Эта мутация нарушает пролиферацию не только меланобластов, но и других производных нервной трубки. Этим объясняется отмеченная еще Дарвиным «загадочная «корреляция между белым цветом шерсти, голубым цветом глаз и глухотой кошек. Такое сильное и раннее повреждение развития у носителей данной мутации приводит к снижению их плодовитости и жизнеспособности.
Это является общим правилом: чем раньше в онтогенезе включается мутантный ген, тем шире спектр его плейотропных проявлений, поскольку даже не-значительное, но раннее повреждение определенной структуры вызывает каскад изменений во всех других структурах, которые от нее происходят или зависят в своем развитии.
Если бы кошки были предоставлены сами себе, то эта мутация давно была бы стерта с лица Земли. Однако восторженное отношение к белым котам со стороны кошковладельцев, а также известное тщеславие («Ни у кого нет такого кота, а у меня есть») приводят к тому, что этот вредный аллель не исчезает из популяции кошки [15].
Можно ожидать, что результатом такого поддерживающего отбора со временем станет уменьшение неблагоприятных плейотропных эффектов аллеля W и повышение плодовитости и жизнеспособности его носителей.
На пути меланобластов от нервной трубки к волосяному фолликулу может встать еще одно препятствие. У животных гомо- и гетерозиготных по доминантной мутации S скорость миграции меланобластов замедлена. Такие клетки не успевают добраться к фолликулам в определенных участках тела до конца их дифференцировки, и эти участки остаются не окрашенными.
Здесь используются понятие генетики - гены-модификаторы. Этим обозначают гены, которые могут тем или иным способом влиять на проявление главного гена, ответственного за определенный признак. Они могут усиливать или ослаблять его эффект. В данном случае гены контролируют либо темп дифференцировки волосяных фолликулов, либо плотность тканей, через которые мигрируют меланобласты. Сочетаясь в разных вариантах они создают различия в генетическом фоне, и общее для всех носителей гена S торможение миграции меланобластов приводит к фенотипически разным результатам (в смысле локализации белого пятна).
В зависимости от набора генов - модификаторов фенотипическое проявление мутации может варьировать от маленького белого пятна на груди до настолько сильной экспансии белых районов, что на их фоне участки нормальной окраски кажутся пятнами.
Когда меланобласты достигли волосяных фолликулов, они входят в них и здесь заканчивают свою дифференцировку. Теперь они называются меланоциты и приступают к синтезу пигментов, черного и желтого. Этот процесс может быть нарушен целым рядом мутаций. Одну из них мы уже рассмотрели (О). Эффект этого гена проявляется только в меланоците.
Другая мутация (а) приводит в гомозиготном состоянии к нарушениям распределения желтого пигмента вдоль по волосу. Носители этой мутации имеют черную окраску меха. Эффект этого гена не проявляется в клетках с генотипом О/О и О/У.
Может быть также нарушен синтез обоих типов пигмента. Фермент тирозиназа играет ключевую роль в процессе превращения тирозина в меланин. Деффект в функции этого фермента обычно является причиной возникновения разных вариантов альбинизма. Предполагают, что таким деффектом обладают сиамские кошки. Они имееют в гомозиготном состоянии ген Сs.
В данном случае функция тирозиназы не нарушена полностью. У сиамских кошек активность этого фермента в сильной степени определяется температурой. При нормальной температуре тела его активность резко снижена, в то время как при пониженной температуре этот фермент оказывается способным к выполнению своей функции. Именно поэтому в тех частях тела, где температура снижена - на ушах, носу, хвосте, конечностях, - формируются интенсивно окрашенные участки, а туловище окрашено значительно слабее.
Это пример взаимодействия генотипа со средой. Если сиамского кота выращивать на холоде, он будет совсем темный, а если прикладывать к его конечностям грелку, он вырастет светлым.
Мутации затрагивают не только синтез пигмента, но также структуру пигментных гранул в волосе. Примером мутации такого типа служит мутация ослабленной окраски - (d). Этот ген контролирует не количество пигментов, а определяет форму меланоцитов. Мутантные меланоциты имеют короткие дендритные отростки и передают незначительное количество пигментных гранул в волос. Гранулы в таком волосе распределяются не равномерно, а образуют локальные скопления. Создается впечатление ослабленной окраски, хотя интенсивность синтеза пигмента нормальная. Эта мутация оказывает существенное влияние на проявление других мутантных генов.
Ген О у животных, гомозиготных по мутации d, дает кремовую окраску, а гомозиготы по генам аа и dd выглядят серовато-голубыми.
По признаку полосатости обнаружены мутации двух типов: доминантная (Т), которая препятствует формированию рисунка на всем теле за исключением головы, конечностей и хвоста, и рецессивная (t), которая в гомозиготном состоянии превращает полосы в разводы причудливой формы.
Разобравшись с самыми распространенными мутациями окраски, можно в известной мере предсказать расцветку будущих котят, зная масть их родителей [15].
Геногеография кошек
Генные частоты описаны в 150 популяциях из разных точек земного шара от Сингапура до Сан-Пауло. Методы сбора материала были самыми разнообразными: это и ветеринарная статистика, и любезная помощь обществ призрения животных, и поквартирный обход, и просто прогулки по городу с блокнотом.
Какие же результаты были получены при картировании частот кошачьих популяций?
Относительно высока частота рыжих кошек в двух разных районах - в сфере влияния арабских калифов в Малой Азии, на южном побережье Средиземного моря и в зоне действия викингов - Британские острова, Бретань, Фландрия. Там же, где некогда простиралась Западная Римская империя, аллель О встречается относительно редко.
Очень интересны в этом отношении исследованные популяции отечественных кошек. Популяции Санкт-Петербурга и Сибири ведут свои родословные, по-видимому, от кошек, что пришли к нам вместе с Рюриком. Здесь аллель О встречается с частотой 20-25%. А вот в Анапе и Краснодаре - в районе, над которым вначале нашей эры простирал свои крыла римский орел, насчитали крайне мало рыжих кошек.
Итак, благодаря геногеографии, мы можем делать различные допущения относительно психологии наших пращуров. Римляне, видимо, не одобряли рыжих кошек, а вот арабы и викинги - напротив…
Частота генов в популяции - это показатель очень устойчивый. В больших популяциях при отсутствии отбора по признаку, контролируемому данным геном, и селективной миграцией он может сохраняться без изменения десятки и сотни поколений. Естественно, что 1500 лет после падения Рима в кошачьих популяциях произошло много изменений, связанных с миграциями людей и кошек. Вместе с тем различия в генных частотах по аллелю О, возникшие, видимо, еще в то время, до сих пор не стерлись.
Распределение аутосомного аллеля Т, который вызывает исчезновение рисунка на теле кошки, тоже заставляет нас вспомнить историю и этнографию. Этот аллель встречается, довольно редко, в относительно узком коридоре - Сингапур, Таиланд, Новосибирск, Куйбышев, Будапешт, Вена. В других районах его почти нет.
Известно, что переселение народов с Урала и Поволжья в конце первого тысячелетия нашей эры привело к формированию современного населения Венгрии. Кошки, как всегда, следовали за людьми и несли свои гены в Европу. С этим и связано сходство в генетических профилях указанных выше популяций, которые находятся довольно далеко друг от друга [15].
До сих пор мы рассматривали распределение доминантных мутантных аллелей в кошачьих популяциях. Однако особенно изящные клинальные карты получены для рецессивных аллелей. Мутация l, которая ответственна за формирование длинного пушистого меха. Родиной этой мутации следует считать север европейской части СНГ, где частота ее максимальна. По мере удаления от центра ареала длинношерсные кошки встречаются все более и более редко.
Можно предполагать, что повышение частоты аллеля l в России вызвано отбором, проводимым человеком. Сибирские коты пользовались у нас всегда большой популярностью. Кроме того, нельзя исключить возможности естественного отбора в пользу длинношерстности. Действительно, частота этого гена на юге резко падает. В Краснодаре он очень редок, а в Анапе гомозиготы по этому аллелю не обнаружены вовсе. Хотя кошки, как правило, живут в теплых помещениях, тем не менее можно предположить, что в районе с более холодным климатом особи с длинной, пушистой шерстью обладают селективным преимуществом перед своими нормальными собратьями.
Распределение другого рецессивного аллеля t находится в Англии и Иране. В этих точках частота мраморных котов максимальная. В Сибирь носители этого гена до сих пор не проникли в частоте, достаточной для выщепления гомозигот по данной мутации.
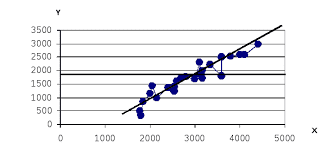
На основании анализа популяций кошек в Англии и ее бывших колониях была выявлена одна очень интересная закономерность в изменениях частот аллеля t. Если построить график зависимости частоты гена t от времени возникновения популяции, легко можно увидеть, что зависимость имеет линейный характер. Чем раньше попали английские кошки в этот или иной район, тем меньше там частота мраморных кошек.
Если продолжить эту линию до наших дней, то она даст значение частоты в современной Англии. На основании данного графика специалист по популяционной генетики кошки Нейл Б. Тодд высказал следующее предположение. В силу непонятных причин частота аллеля t в Англии очень быстро растет, в то время как в ее бывших колониях она остается неизменной. Тогда генетические профили популяций бывших колоний можно рассматривать как исторические портреты старинных английских популяций [16].
Ген а можно назвать аллелем урбанизации. Его частота возрастает прямо пропорционально росту численности населения.
Почему же в городах черные кошки встречаются чаще, чем в деревнях? Подозревать сельских жителей в том, что они более суеверны, нет оснований. Доктор Кларк из Шотландии провел в Глазго и его пригородах опрос населения. Он спрашивал: «Как вы думаете, черный кот приносит счастье, несчастье или вообще не влияет на фортуну?» И в городе и в деревне ответы были одинаковыми: 20% уверены, что он приносит счастье, 20% - что несчастье, 60% - не отнеслись к этому вопросу всерьез.
Таким образом, гипотеза об искусственном отборе на основе суеверия отпадает. Может быть, здесь действует естественный отбор? Не является ли наблюдаемый нами феномен еще одним примером промышленного механизма?
Меланисты (животные с черной окраской) оказываются более спокойными, более дружелюбными к человеку, более устойчивыми к действию стрессов [17]. Таким образом, можно предположить, что в силу особенностей своего поведения черные кошки более приспособлены к напряженной и нервной городской жизни, чем их нормальные сородичи. Именно этим и объясняется резкое возрастание частоты аллеля а в городских популяциях.
Теперь можно убедиться какие интересные и важные выводы нужно сделать из результатов изучения геногеографии кошек [18].
Материалы и методы
Основным методом исследования является визуальное типирование характера и окраски шерстного покрова животных, встреченных на улицах, во дворах домов, в парках, скверах и т.п.
Частоты рецессивных аллелей (q) определялись путем извлечения квадратичного корня из частот соответствующих мутантных фенотипов, а доминанных (p) - из соотношения:
p= 1 - q.
Ошибки частот подсчитывались по формулам для рецессивных и доминантных аллелей соответственно, где n - общее число исследованных животных, определенное для каждого локуса с учетом того, что аллель W эпистатичен относительно всех аллелей, влияющих на окраску аллель а - относительно аллелей Т и t, аллель О - относительно аллеля а [20].
Доля самцов (m) определялась из соотношения m=(4аc-b)/(2a+b) (b+2c) [23]. Для получения ожидаемого соотношения фенотипов локуса удовлетворяющего равенству m = 0.5, ожидаемое число оранжевых животных принималось равным наблюдаемому; ожидаемое число черепаховых животных вычислялось из соотношения: b = n (8a+n) - (2a+n); ожидаемое число неоранжевых - из соотношения c = n - (а + b) [24].