2020-05-12
2020-05-12 396
396Промоторы (к ним присоединяются РНК – полимераза, чтобы начать транскрипцию).
Терминаторы (на таких участках РНК – полимераза заканчивает транскрипцию).
Операторы (к ним присоединяются белки – репрессоры, включающие работу РНК - полимеразы).
Энхансеры (усиливать) участки ДНК, к которым присоединяются особые белки, изменяющие скорость транскрипции и тем самым скорость синтеза соотносимых белком.
Сайленсеры (заглушать) или это последовательность ДНК, расположенные в тысячах пар нуклеотидов от промотора эукариотического гена и оказывающие дистанционное влияние на его транскрипцию.
3. По влиянию на физиологические процессы клетки
• летальные,
• полулетальные,
• мутаторы,
• антимутаторы,
• гомеобоксные.
Отделы ДНК:
Между генами находится некодирующие последовательности - СПЕЙСЕРЫ
Функциональная роль спейсеров:
- Выполняют структурную роль:
• Участвуют в правильной укладке нуклеосомной цепи в высшие структуры хроматина;
• В прикреплении хромосом к аппарату центриолей.
2. Служат специфическими локусами связывания определенных белков:
• функционирующих на ДНК ферментов (ДНК – полимеразного комплекса)
• белков, выполняющих регуляторную функцию
При этом участки связывания РНК-полимеразы (фермента, синтезирующего РНК на ДНК) называются промоторами. Они либо вплотную примыкают к началу гена (или группы генов), либо отделены от гена какими-либо другими функциональными локусами. Характерный компонент промоторов у кишечной палочки – бокс Прибнова:
- ТАТААТ -
- АТАТТА -
Он находится за 15 н.п. от стартовой точки транскрипции. Общая же протяженность промотора – несколько десятков н.п.
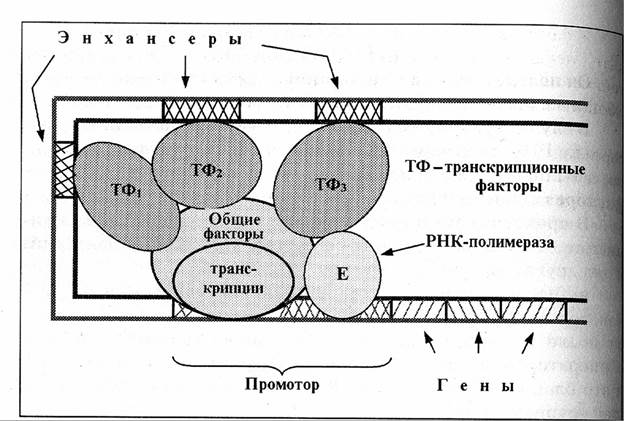
В случае эукариот промотор – более сложное понятие, поскольку РНК-полимераза связывается с ДНК не непосредственно, а лишь вместе с комплексом других белков – т.н. общих факторов транскрипции. В промоторе эукариот различают небольшую область инициации, ТАТА-бокс (сходный с боксом Прибнова у бактерий) и ряд других типичных участков. У эукариот регуляцию «прочтения» генов осуществляют не только белки-репрессоры, но и белки-активаторы – т.н. транскрипционные факторы. К последним относятся уже упоминавшиеся общие факторы транскрипции, необходимые для связывания РНК-полимеразы с промотором. Эти факторы имеются во всех клетках и необходимы для «прочтения» любого функционирующего гена.
Другие транскрипционные факторы повышают активность только определенных генов, и локусы ДНК, связывающие такие факторы, называются энхансерами. Энхансеры могут располагаться достаточно далеко от регулируемого гена: на расстоянии нескольких тысяч нуклеотидных пар.
Как же связывание с ними транскрипционных факторов может стимулировать активность гена?
Видимо, дело в сложной пространственной организации ДНК. Последняя образует петли, благодаря чему энхансеры сближаются с промоторной зоной и связанные с ними факторы влияют на активность транскрипционного комплекса.
Причем, для некоторых ключевых генов в клетке имеется сразу несколько энхансеров, достаточно удаленных друг от друга. Следовательно, все они в результате изгибов ДНК должны собираться примерно в одном месте пространства.
Энхансеры - усилители транскрипции – обладают следующими свойствами:
- могут находиться как в 5, так и в 3 – областях, а также в интронах и даже на значительном расстоянии от промоторов;
- активируют гены независимо от ориентации;
- один энхансер может активировать различные гены;
- действие их может быть ткане- и видоспецифичным;
- энхансеры доступны действию различных белков, в том числе и гормонов.
Сайленсеры - ослабители транскрипции – являются негативными элементами по отношению к транскрипции. Они так же, как и энхансеры могут оказывать свое действие на большом расстоянии от гена и при разной ориентации по отношению к нему.
3. В ДНК могут содержаться короткие локусы, служащие сигналами об окончании транскрипции ДНК (терминаторы). У бактерий в ряде случаев участки с такой функцией находятся перед группой совместно регулируемых генов. Это т.н. аттенюаторы. Терминирующие же участки, располагающиеся после генов, называются терминаторами. В одних условиях транскрипция прекращается на аттенюаторе (гены не считываются), а в других условиях – на терминаторе (гены прочитываются).

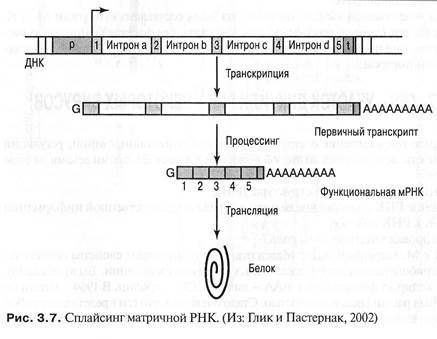
Мозаичную структуру эукариотных генов открыли в 1977 году группой ученых, возглавляемых американскими исследователями Ричардом Робертсом и Филиппом Шарпом.
Эукариотические гены имеют мозаичную структуру и состоят из кодирующих участков – экзонов и расположенных между ними некодирующих областей – интронов. При транскрипции ДНК считывается целиком, а затем образовавшаяся пре-мРНК подвергается созреванию (процессингу): участки РНК транскрибированные с интронов, вырезаются, а участки РНК, синтезированные на экзонах, сшиваются (сплайсинг)