 2020-05-21
2020-05-21 522
522Сцепленные гены - гены, находящиеся в одной хромосоме.
Группа сцепления – все гены, расположенные в одной хромосоме. Все гены, относящиеся к одной группе сцепления, при образовании гамет наследуются вместе.
Сцепленное наследование генов – совместное наследование генов, локализованных в одной хромосоме.
Рассмотрим опыт Моргана.
Объект: мухи дрозофилы.
Вид эксперимента – дигибридное скрещивание.
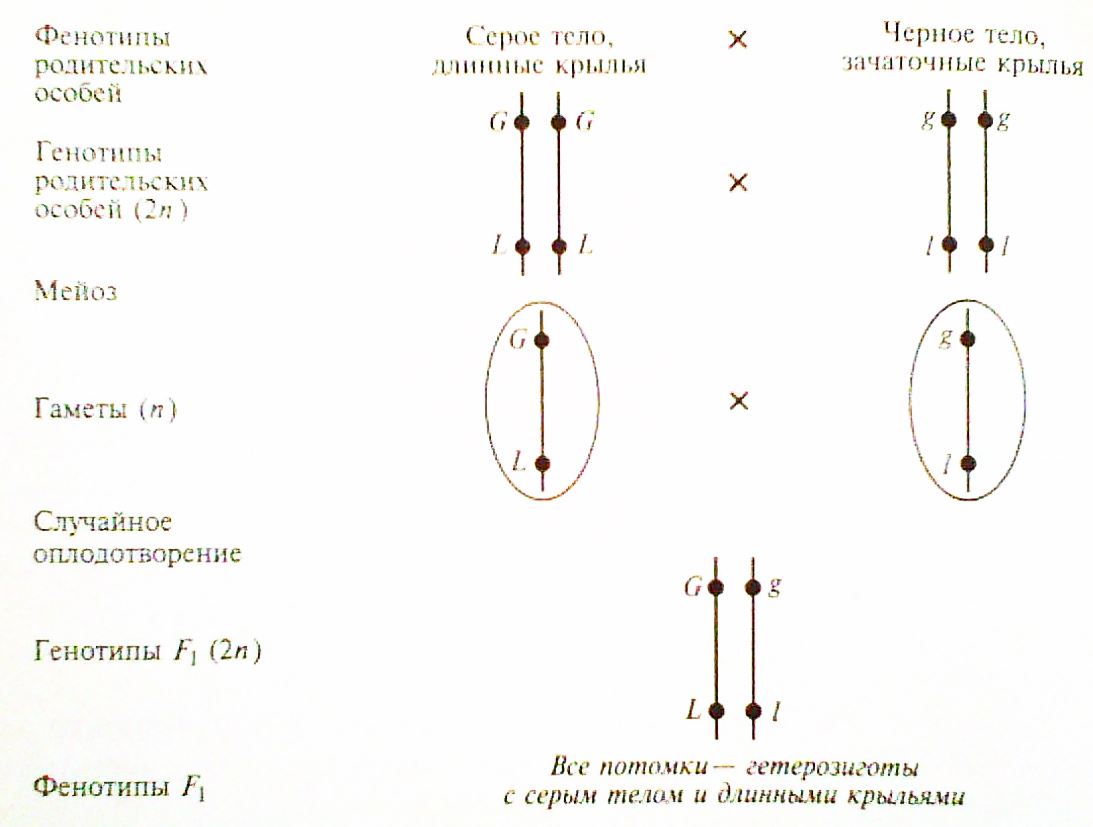
Условия: скрестили мух, имеющих серое тело и нормальные крылья, с мухами, имеющими темное тело и укороченные крылья.
G – ген, определяющий серое тело, g – ген, определяющий темное тело;
L- ген, определяющий нормальную длину крыльев, l – ген, определяющий короткие крылья.
Эксперимент 1. Родители – доминантные и рецессивные дигомозиготы.
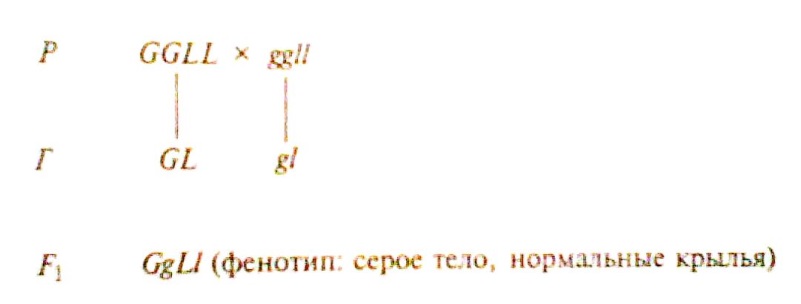
(генотип: дигетерозиготы)
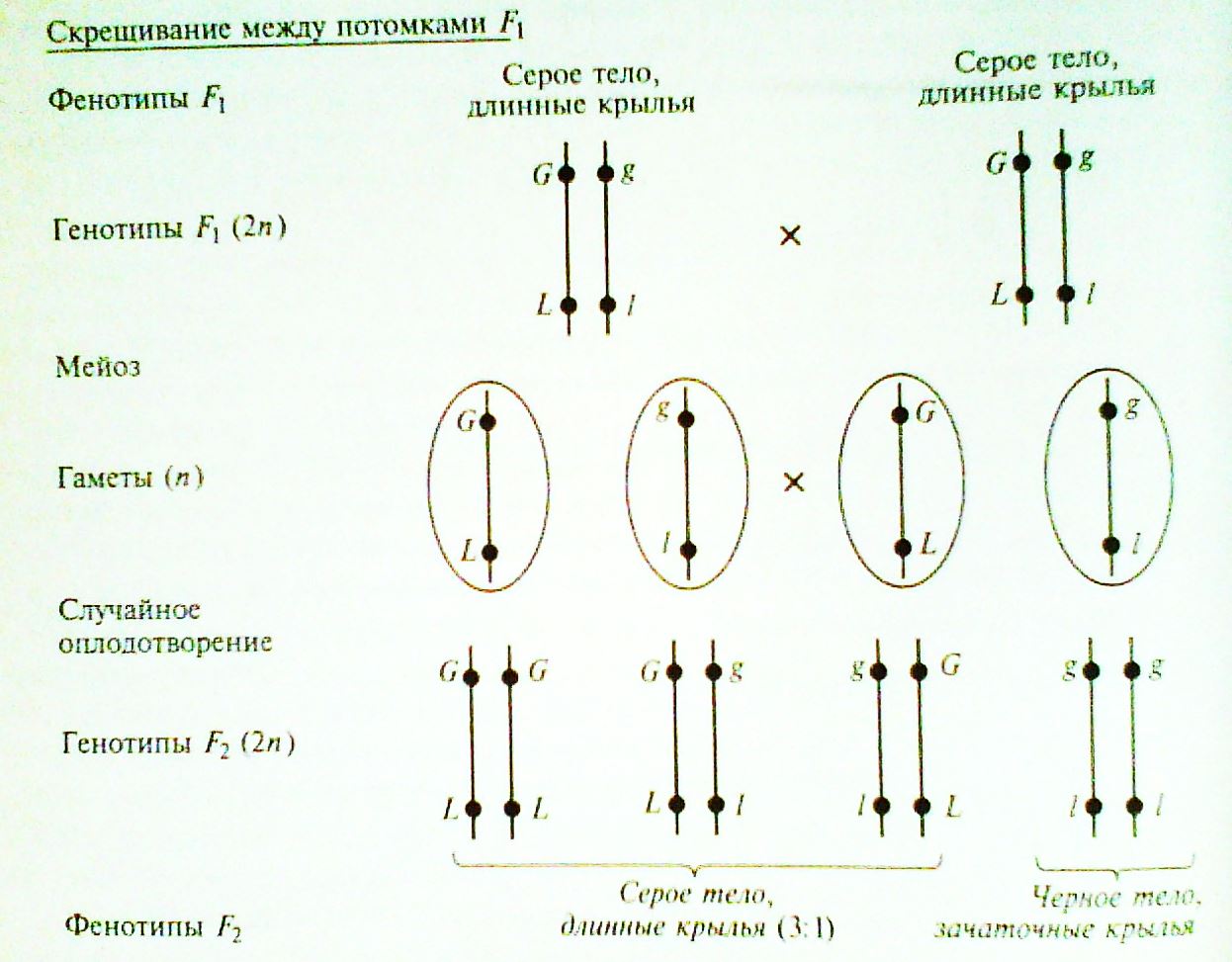
Скрестив гибридов первого поколения между собой, получили расщепление 3:1, как при моногибридном скрещивании (по фенотипу 3 с серым телом и нормальными крыльями: 1 с темным телом и короткими крыльями)
Цитологигические основы эксперимента (были установлены позже):
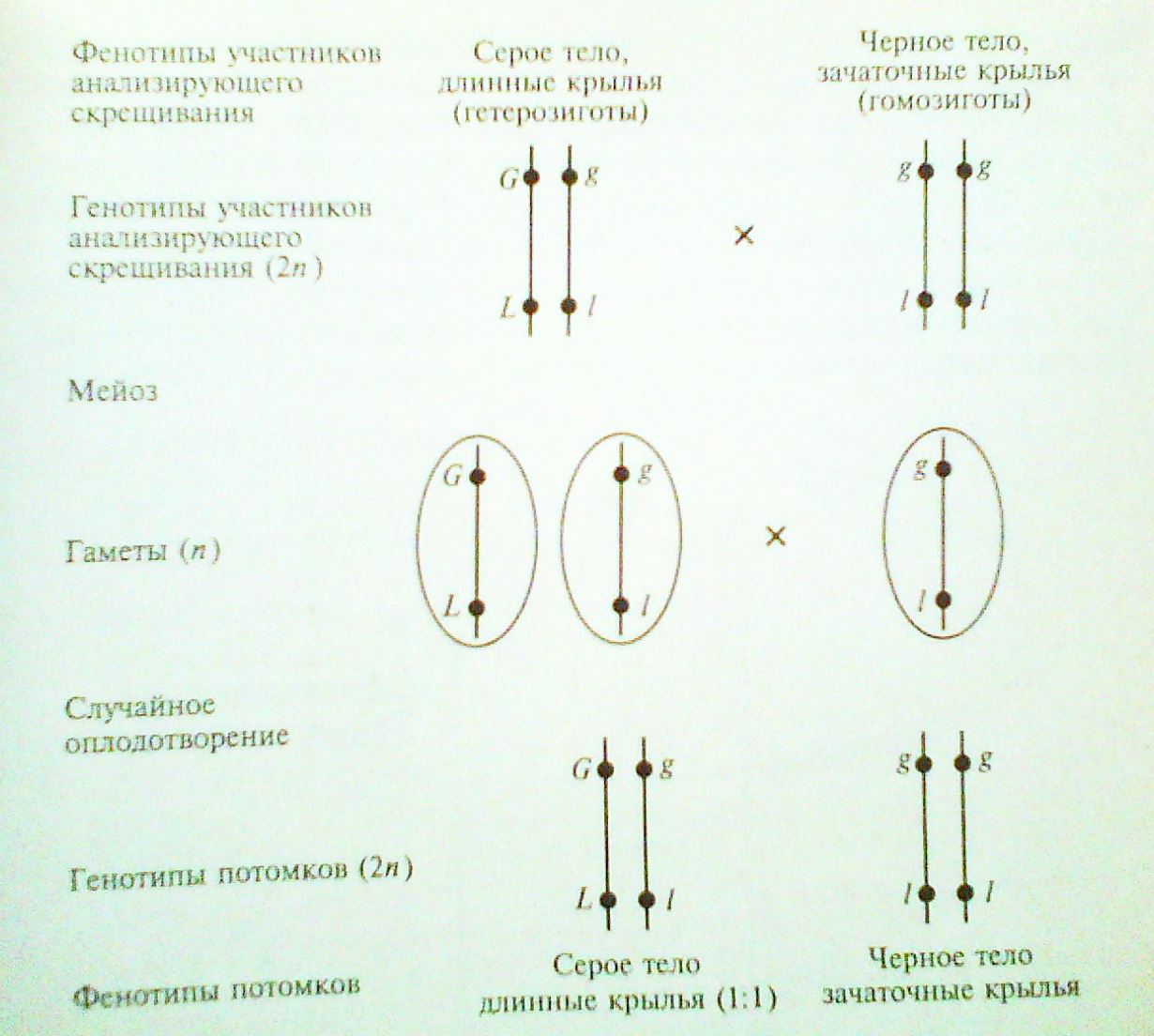
Эксперимент 2. Провели дигибридное анализирующее скрещивание гибридов первого поколения с эталонными образцами (рецессивными дигомозиготами).
Ожидаемые результаты:
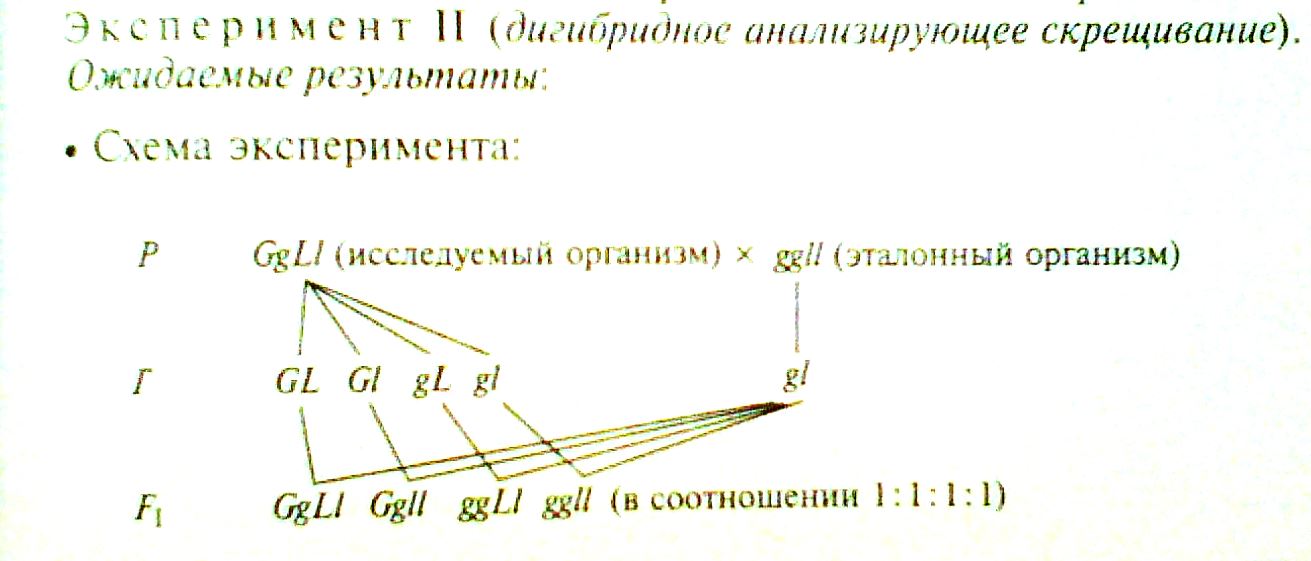
Предполагалось, что по фенотипу получится расщепление
1 серое тело, нормальные крылья: 1 серое тело, короткие крылья: 1 темное тело, нормальные крылья: 1 темное тело, короткие крылья,
а по генотипу 1дигетерозигота: 1 гетерозигота по цвету тела, рецессивная гомозигота по длине крыльев: 1 гетерозигота по длине крыльев и рецессивная гомозигота по цвету тела: 1 рецессивная дигомозигота.
Фактический результат оказался другим: F1 GgLl: ggll (в соотношении 1:1)
Отсутствие в потомстве особей GgLl и ggLl указывает на отсутствие гамет типа Gl и gL. Причина: гены G и L, так же, как и гены g и l, располагаются в одной хромосоме, то есть сцеплены.
Цитологическая основа эксперимента:
Так был открыт закон: гены, расположенные в одной хромосоме, наследуются вместе и составляют группу сцепления (закон Моргана).
Количество генов в различных группах сцепления (то есть в различных хромосомах) может отличаться друг от друга. При дигибридном скрещивании сцепленные гены, как правило, не подчиняются законам Менделя. С другой стороны, полное сцепление случается достаточно редко, и в потомстве обычно бывают представлены все четыре фенотипа. Таким образом, и в этом случае при дигибридном скрещивании образуются новые сочетания признаков – рекомбинантные фенотипы. Итак, если особи с новыми генными комбинациями встречаются в потомстве реже, чем особи с родительскими фенотипами, то это верный признак сцепленности соответствующих генов.
При многократном повторении эксперимента № 1 среди потомства в небольшом проценте случаев стали встречаться особи, «находящиеся под запретом» - GgLl и ggLl. Тщательная проверка данного феномена на других объектах подтвердила достоверность полученных фактов. Для их объяснения была предложена концепция неполного сцепления генов. Отрицающая жесткое сцепление генов в хромосоме и допускающая возможность обмена соответствующими локусами между гомологичными хромосомами при кроссинговере. Появление рекомбинантных сочетаний у аллелей называется кроссинговером ( перекрестом ). Именно таким образом возникают гаметы Gl и gL и особи GgLl и ggLl, получившие название кроссоверных.
При дальнейшем изучении этого явления была выявлена закономерность: частота появления кроссоверных гамет прямо пропорциональна расстоянию между соответствующими генами в хромосоме. Дистанцию между генами принято выражать в процентах или в условных единицах – морганидах.
Цитологическая основа неполного сцепления генов:
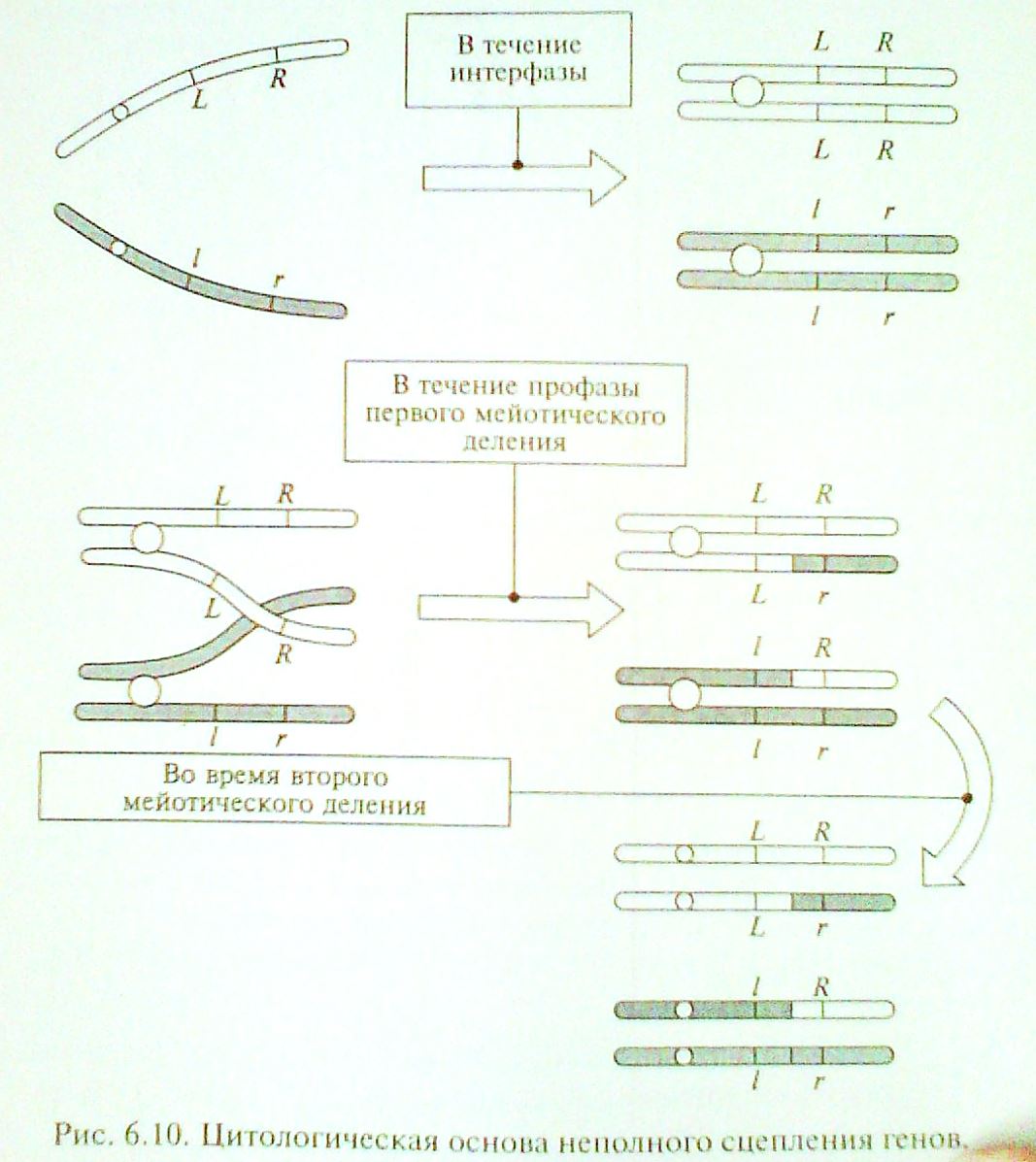
Исследования генетиков начала XX века показали, что кроссинговер имеет место в результате разрыва и рекомбинации гомологичных хромосом и происходит практически между всеми хромосомами. Частота рекомбинаций определяется по формуле
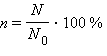 , где N – количество рекомбинантов, N0 - общее количество потомков.
, где N – количество рекомбинантов, N0 - общее количество потомков.
В то же время частота рекомбинаций определяет число рекомбинаций, происходящих при образовании гамет.
Частота рекомбинаций генов показывает относительное расположение сцепленных генов в хромосоме: чем дальше друг от друга находятся гены, тем выше частота рекомбинации. Это обстоятельство используется при составлении генетических карт. Условное «расстояние» между локусами (местоположениями в хромосоме) двух генов считается прямо пропорциональным частоте рекомбинации. Взаимное расположение (последовательность) локусов трёх и более генов определяется методом триангуляции. При этом сначала берутся гены с наименьшей частотой рекомбинации. Далее выбирают следующую по величине частоту рекомбинации и указывают два возможных положения нового гена; одно из этих положений будет отсеяно на следующем шаге, когда берётся третья частота. В реальных экспериментах генетические карты могут искажаться благодаря двойному кроссинговеру, когда рекомбинация происходит одновременно в двух точках. Двойной кроссинговер особенно характерен для генов, локусы которых разделены большими расстояниями.
Хромосомная теория наследственности (предложена Т. Морганом в 1911-1926 годах)
1. Гены локализованы в хромосомах. Набор генов каждой из негомологичных хромосом уникален.
2. Гены расположены в хромосоме линейно.
3. Аллельные гены занимают одинаковые локусы в гомологичных хромосомах.
4. гены одной хромосомы образуют группу сцепления.
5. Сцепление неполное, возможен кроссинговер.
6. Для каждого биологического вида характерно свое число групп сцепления.








