2020-05-25
2020-05-25 336
336Водно-солевой гомеостазис является одной из важнейших констант внутренней среды организма. От состояния водно-солевого обмена зависят фундаментальные свойства клеток и тканей (энергетический и пластический метаболизм, возбудимость, проводимость и сократимость, секреторная способность и т. п.) и, соответственно, функции практически всех физиологических систем. Основные характеристики водно-солевого обмена — количество поступающих в организм жидкости и солей, объемы внеклеточных и внутриклеточных водных пространств, содержание в них отдельных электролитов и осмотическое давление, наконец, выделение воды и солей почками — имеют взаимосвязанные комплексные механизмы регуляции. Эти механизмы легко проследить на примерах изменения приема жидкости или ее потерь, сдвигов баланса ионов натрия и калия, изменения объема крови.
Нарушения баланса веществ могут проявляться в виде положительного или отрицательного дисбаланса. Положительный водный дисбаланс получил название гипергидратации, а отрицательный — дегидратации. Изменения водного баланса могут сопровождаться или не сопровождаться изменениями электролитного баланса и, соответственно, осмотического давления. В зависимости от осмолярности виды водного дисбаланса подразделяют на изоосмотические, гиперосмотические и гипоосмотические. Эти понятия относят к внеклеточной среде организма, но благодаря механизмам саморегуляции поддерживается осмотическое равновесие между вне- и внутриклеточной средой, поэтому изменения осмолярности внеклеточной среды неизбежно сопровождаются аналогичными сдвигами осмотического давления и внутри клеток. Водный дисбаланс может быть и следствием изменения коллоидно-осмотического (онкотического) давления плазмы крови.
14.4.1. Гомеостатические механизмы при гиперосмотической дегидратации
Такой вид водного дисбаланса возникает при дефиците поступления воды в организм, а также в случаях, когда потери воды превышают потери солей или при потерях свободной от электролитов воды. Примером этого типа дисбаланса является пребывание человека в пустыне, когда недостаток воды для питья сочетается с повышенными перспирационными (через кожу и дыхательные пути) потерями воды. Недостаточное поступление воды в организм может быть следствием отсутствия чувства жажды при патологии диэнцефальной области головного мозга, нарушений режима кормления и ухода за детьми грудного возраста, больными при потере сознания, нарушений глотания.
Ограничение приема воды (рис. 14.14) приводит к дегидратации внутренней среды организма, при этом имеет место повышение осмотического давления плазмы крови и внеклеточной жидкости, что дало основание именовать этот вид дегидратации гиперосмотической, или истинной. Повышение осмотического давления плазмы крови вызывает перераспределение жидкости и электролитов между кровью, внеклеточной жидкостью и лимфой, а также между внеклеточной и внутриклеточной средой, направленное на нормализацию сдвига осмотического давления. Начальные механизмы регуляции гомеостазиса при этом типе дегидратации заключаются в буферировании белками и эритроцитами катионов, образовании с ними сложных химических соединений, с измененными осмотическими свойствами и гидрофильностью, диффузии осмотически активных веществ из крови в тканевую жидкость и лимфу, осмотическом токе воды в противоположном направлении, усилении всасывания воды в кишечнике. При небольших физиологических уровнях дегидратации этих механизмов может быть достаточно для обеспечения водно-солевого гомеостазиса.
Повышенное осмотическое давление жидкостей внутренней среды раздражает осморецепторы, приводя к формированию осморегулирующих рефлексов и жажды.
Осморегулирующие рефлексы начинаются от периферических и центральных осморецепторов (рис. 14.15). Периферические осморецепторы расположены в интерстициальном пространстве тканей и кровеносных сосудах печени (система воротной вены), сердца, пищеварительного тракта, почек, селезенки, а также в рефлексогенной зоне каротидного синуса. Поскольку осмотическое давление связано преимущественно с ионами натрия, часть осморецепторов являются специализированными натриорецепторами. Наиболее мощными рецептивными полями для натрия являются интерстициальные пространства печени и предсердий. Имеются также тканевые рецепторы, воспринимающие концентрацию ионов калия, кальция и магния. Центральные осморецепторы — клетки супраоптического ядра гипоталамуса, непосредственно воспринимающие сдвиги осмотического давления притекающей в межуточный мозг крови и концентрацию в ней натрия (натриорецепторы).
Афферентная информация от тканевых осмо- и натриорецепторов поступает по волокнам блуждающего нерва и задних корешков спинного мозга в центральную нервную систему, где направляется в центр осморегу-
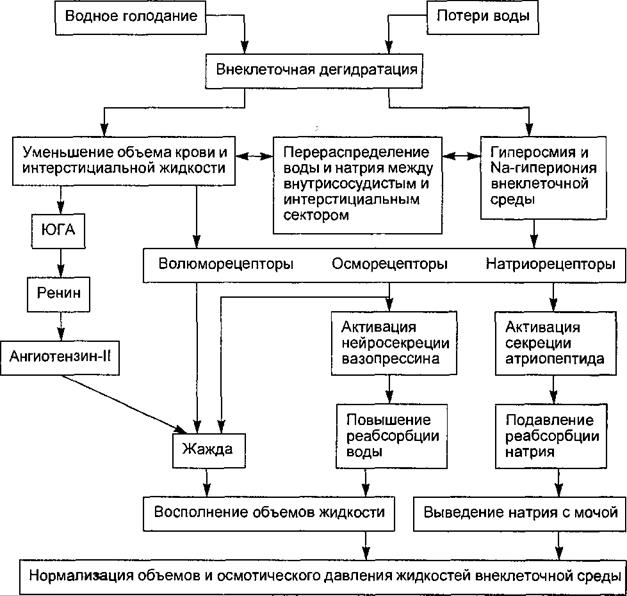
Рис. 14.14. Схема компенсаторных реакций при ограничении приема или потерях воды, т. е. при гиперосмотической дегидратации. Основные реакции направлены на восполнение объемов воды (жажда, повышение реабсорбции воды в почках) и выведение избытка натрия с мочой (угнетение реабсорбции катиона в почках).
ляции, локализованный в ядрах переднего гипоталамуса — супраоптическом и паравентрикулярном. Эфферентное звено осморегулирующих и на- трийрегулирующих рефлексов включает как вегетативные нервы, так и гормональные сигналы. Осмо- и натрийрегулирующие рефлексы увеличивают нейросекрецию и содержание в крови вазопрессина. Избыток ионов натрия в крови стимулирует секрецию натрийуретического атриопептида. Вазопрессин повышает в почках проницаемость дистальных канальцев и собирательных трубочек для воды, и она пассивно реабсорбируется по осмотическому градиенту, т. е. реализуется механизм концентрирования мочи. Вазопрессин повышает всасывание воды и в других органах — кишечнике, желчном пузыре, слюнных и пищеварительных железах. Натрийуретический атриопептид, как и, в меньшей степени, вазопрессин, ведут к повышенному выведению натрия из крови, что устраняет увеличение осмотического давления.
 |
|
 Активация реабсорбции воды, концентрирование мочи, снижение осмолярности крови
Активация реабсорбции воды, концентрирование мочи, снижение осмолярности крови