 2020-06-10
2020-06-10 4811
4811Вопросы к теме:
1. Гипотезы происхождения жизни. Изучение основных закономерностей возникновения, развития и существования жизни на Земле.
2. Усложнение живых организмов в процессе эволюции. Многообразие живого мира на Земле и современная его организация.
1. Гипотезы происхождения жизни. Изучение основных закономерностей возникновения, развития и существования жизни на Земле:
Гипотезы происхождения жизни:
Происхождение жизни на Земле является одной из важнейших проблем естествознания. На протяжении десятков веков менялись взгляды на проблему жизни, высказывались разные идеи, гипотезы и концепции. Некоторые из них получили широкое распространение в разные периоды истории развития естествознания. В настоящее время существует пять гипотез возникновения жизни:
1. Креационизм – гипотеза, утверждающая, что жизнь создана сверхъестественным существом в результате акта творения. Имеет самую длинную историю. Основывается на наличии в живых организмах особой силы, «души», которая управляет всеми жизненными процессами.
2. Гипотеза стационарного состояния, согласно которой жизнь никогда не возникала, а существовала всегда. С изменением природных условий изменялись и виды: одни исчезали, другие появлялись. Основывается на исследовании палеонтологов.
3. Гипотеза самопроизвольного зарождения жизни, которая основывается на идее многократного возникновения жизни из неживого вещества, была выдвинута в древнем Китае и Индии как альтернатива креационизму. Эту гипотезу поддерживали Платон, Аристотель, Галилей, Декарт, Ламарк. Суть гипотезы: низшие живые организмы возникли из ила, сырой почвы, гниющего мяса. В опровержение этой гипотезы Ф.Реди сформулировал принцип: «Все живое – от живого», после того, как нашел причину появления червей на гниющем мясе. Л.Пастер своими опытами с вирусами окончательно доказал несостоятельность идеи спонтанного зарождения жизни.
4. Гипотеза панспермии, согласно которой жизнь была занесена на Землю из космического пространства. Впервые была высказана Г.Рихтером в конце 19 столетия. Данная концепция допускает возможность происхождения жизни в разное время в разных частях Вселенной и переносе ее различными путями на Землю (метеориты, астероиды, космическая пыль).
5. Гипотеза исторического происхождения жизни путем биохимической эволюции. Авторами являются А.Опарин и С.Холдейн. С точки зрения гипотезы А.Опарина, а также с позиций современной науки возникновение жизни из неживого вещества произошло в результате естественных процессов во Вселенной при длительной эволюции материи. А.Опарин выделил несколько этапов биохимической эволюции, конечной целью которых явилась примитивная живая клетка. Эволюция шла по схеме:
А) геохимическая эволюция планеты Земля, синтез простейших соединений, таких как СО2, NH3, H2O и т.д., переход воды из парообразного состояния в жидкое в результате постепенного охлаждения Земли. Эволюция атмосферы и гидросферы.
Б) образование из неорганических соединений органических веществ – аминокислот – и их накопление в первичном океане в результате электромагнитного воздействия Солнца, космического излучения и электрических разрядов.
В) постепенное усложнение органических соединений и образование белковых структур.
Г) выделение белковых структур из среды, образование водных комплексов и создание вокруг белков водной оболочки.
Д) слияние таких комплексов и образование коацерватов, способных обмениваться веществом и энергией с окружающей средой.
Е) поглощение коацерватами металлов, что привело к образованию ферментов, ускоряющих биохимические процессы.
Ж) образование гидрофобных липидных границ между коацерватами и внешней средой, что привело к образованию полупроницаемых мембран, которые обеспечивали сохранение стабильности функционирования коацервата.
З) выработка в ходе эволюции у этих образований процессов саморегуляции и самовоспроизведения.
По мнению академика В.Вернадского возникновение жизни связано с мощным скачком, который внес в эволюцию столько противоречий, что они создали условия для зарождения живой материи. Чрезвычайная сложность организации живой материи является доказательством того, что зарождение жизни является результатом длительного процесса биологической эволюции.
Изучение основных закономерностей возникновения, развития и существования жизни на Земле:
Существуют две альтернативные точки зрения на вопрос о происхождении жизни на Земле:
· теория абиогенеза (живое развивается из неживого)
· теория биогенеза.
Согласно теории биогенеза, живое произошло от живого. Эта теория отрицает самопроизвольное зарождение жизни и подразумевает, что жизнь могла возникнуть, когда возникла неживая материя.
История развития взглядов на происхождение жизни.
Идеи происхождения живого из неживого были распространены в Древнем Китае, Вавилоне, Древнем Египте и Древней Греции.
Выдающийся древнегреческий ученый и мыслитель Аристотель допускал возможность самозарождения организмов. В сочинении «О частях животных» он высказывал идею постепенного перехода от неживых тел к растениям, а от растений к животным. Ученый допускал нелепые, с современных позиций, предположения о том, что черви возникли из ила, клопы — из соков тела животных и т.д. Благодаря тому что взгляды Аристотеля были догматизированы средневековой церковью, идея самозарождения жизни продолжала господствовать и развиваться в Средние века.

Аристотель (384-322 до н.э.)
В 1661 г. итальянский естествоиспытатель и врач Ф. Реди (1626 — 1698) проделал серию оригинальных опытов. Он накрывал сосуды с мясом кисеей, чтобы на него не могли садиться мухи. На его поверхности так и не появились личинки мясных мух. Отсюда Реди сделал вывод, что личинки появлялись не сами по себе, а из отложенных мухами яиц. Это доказывало невозможность самозарождения насекомых.

Франческо Реди (1626-1698)

Опыт Реди
Изобретение в XVII в. голландским натуралистом Антони ван Левенгуком (1632— 1723) микроскопа значительно расширило возможности ученых в изучении строения организмов.

Антони ван Левенгук (1632-1723)
Левенгук изготовлял маленькие, с одной линзой, микроскопы и рассматривал в них различные объекты.
Он впервые увидел бактерии, простейших. Микроорганизмы были обнаружены в самых различных местах, они словно возникали из ничего. Все это, казалось бы, подкрепляло идею самозарождения жизни.
В этом смысле весьма показательными были эксперименты, проведенные в 1748 г. Аббатом и натуралистом из Ирландии Дж. Нидхемом. Исследователь прокипятил мясо в закрытом сосуде, полагая, что высокая температура убьет все зародыши. Но несколько дней спустя в мясном бульоне были обнаружены микроорганизмы. Для ученых того времени это послужило доказательством зарождения живого из неживой материи. Опыты Нидхема повторили многие ученые, и результат всегда был один и тот же. Против выступил итальянский аббат, ученый-натуралист Л.Спалланцани (1729—1799), известный также своими экспериментами по изучению ориентации летучих мышей. Спалланцани проделал тот же опыт, но после кипячения запаял колбу. В этих условиях микроорганизмы не появились в сосуде. Однако виталисты заявили, что длительное кипячение убило «жизненную силу», а в запаянную колбу она не смогла проникнуть.
Даже строгие научные опыты немецкого химика-органика Ф. Велера (1800—1882) по синтезу мочевины (1828) из неорганических соединений, казалось бы, полностью подтверждали идею самозарождения жизни. Идею самозарождения допускал дед Ч.Дарвина — естествоиспытатель-эволюционист Э.Дарвин.

Ф.Велер (1800-1882)

Эразм Дарвин (1731-1802)
Окончательно теория самозарождения организмов была развенчена блестящими работами французского микробиолога Луи Пастера (1822—1895).

Луи Пастер (1822-1895)
В 1862 г. Л. Пастер получил премию французской Академии наук за серию исследований, которыми ученый доказал, что брожение и гниение вызываются бактериями, а не химическими агентами. Данный факт неоспоримо доказывал, что живое возникает только из живого (теория биогенеза).
Опыты Пастера были просты и гениальны. Прокипятив сосуды с питательными средами, в результате чего погибли бактерии и их споры, Пастер, помня о доводах виталистов, не стал запаивать сосуды, а соединил их с наружной средой длинными извитыми трубками. Проникшие сюда споры оседали на стенках длинных изогнутых трубок и не заражали питательную среду, которая оставалась стерильной.
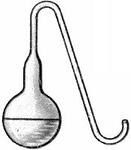
Колба Пастера
Стало абсолютно ясно, что никакой «жизненной силы» не существует.
Широкое хождение в научной среде имела концепция креационизма.
Приверженцы этой теории креационисты считали, что все живые организмы появились на Земле в результате акта творения некоего высшего существа.
Взглядов о постоянстве видов придерживался К.Линней.
А знаменитый французский палеонтолог и сравнительный анатом Жорж Кювье (1769—1832), основоположник теории катастроф (1812), полагал, что отличия флоры и фауны различных геологических эпох — это следствие катастроф, в результате которых вся жизнь погибала, и последующих актов творения, когда создавались новые виды растений и животных.
Ученик Кювье А. Д'Орбиньи насчитал в истории Земли 27 катастроф, после которых в результате божественных актов творения возникали новые виды животных. К середине XIX века благодаря развитию эволюционного учения Ч.Дарвина (1859) теория катастроф сдала свои позиции и утратила былое значение, отойдя в область философии и теологии. Однако в 1864 г. она была возрождена в форме неокатастрофизма австрийским геологом Э.Зюссом (1831 — 1914), который считал, что виды долгое время существуют в стабильном состоянии, а потом при резких изменениях климата интенсивно преобразуются.

Эдуард Зюсс (1831-1914)
Итак, теперь, когда стало ясно, что живое возникает только из живого, предстояло решить главный вопрос: «Как возник самый первый живой организм или зачаток жизни?».
Гипотеза панспермии. В 1865 г. немецкий ученый Г.Рихтер (1818 — 1876) предложил гипотезу панспермии. Окончательно она была сформулирована в 1895 г. шведским ученым С.Аррениусом (1859—1927). Согласно гипотезе, жизнь могла быть занесена на Землю из космоса с метеоритами и космической пылью. Это предположение основывается на данных о высокой устойчивости спор бактерий к радиации, высокому вакууму, низким (близким к абсолютному нулю) и высоким температурам. И тем не менее эта гипотеза не отвечает на главный вопрос, а только переносит место главного действия.

Сванте Аррениус (1859-1927)
Гипотеза биохимической эволюции. В 1924 г. отечественным биохимиком А.И.Опариным (1894— 1980), а спустя 5 лет английским биохимиком и генетиком Дж.Холдейном (1892— 1964) была сформулирована гипотеза, рассматривающая жизнь как результат длительной эволюции углеродных соединений.

Александр Иванович Опарин (1894-1980)

Джон Холдейн (1892-1964)
Согласно гипотезе химической эволюции, изложенной А. И. Опариным в монографии «Происхождение жизни», жизнь, по-видимому, возникла на границе моря, суши и воздуха. Примерно 4 —4,5 млрд лет назад в атмосфере молодой Земли, состоящей из аммиака, метана и паров углекислоты, под действием мощных электрических разрядов могли возникнуть простейшие органические соединения. В растворах белков и нуклеиновых кислот, в так называемом «первичном бульоне», могли возникнуть своеобразные сгустки химических соединений, названные коацерватами.
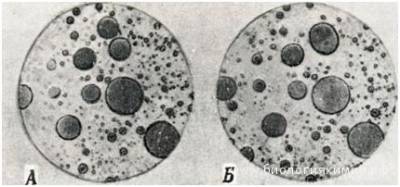
Коацерваты
Несмотря на то что коацерваты способны адсорбировать различные вещества, расти и обмениваться веществами с окружающей средой, их еще нельзя считать живыми существами. Возникновение коацерватов рассматривают обычно как стадию преджизни.
В дальнейшем в результате длительного отбора возникли сложные ферментативные системы, контролирующие процессы синтеза, что обеспечило устойчивость всей структуры. Таким образом, сформировались сложные комплексы нуклеиновых кислот и белков. Нуклеиновые кислоты, способные к воспроизведению, стали контролировать синтез белков, определяя в них порядок аминокислот. В результате сформировался механизм воспроизведения себе подобных и наследования свойств. Так возникло главное свойство живого вещества — способность к воспроизведению подобных себе молекул. Предполагается, что первые организмы были анаэробными гетеротрофами, т.е. получали энергию путем бескислородного расщепления органических соединений. В то время на Земле еще не было свободного кислорода.
Современная теория возникновения жизни на Земле, называемая теорией биопоэза, была сформулирована в 1947 г. английским физиком Дж. Берналом (1901 — 1971).

Джон Десмонд Бернал (1901-1971)
Процесс становления жизни условно разделяют на четыре этапа:
синтез низкомолекулярных органических соединений (биологических мономеров) из газов первичной атмосферы;
образование биологических полимеров;
формирование систем органических веществ, отделенных от внешней среды мембранами (пробионтов);
возникновение простейших клеток, обладающих свойствами живого, в том числе репродуктивным аппаратом, обеспечивающим передачу дочерним клеткам свойств клеток родительских.
Первые три этапа относят к периоду химической эволюции, а с четвертого начинается эволюция биологическая.
Согласно современным представлениям, возраст Земли составляет 4,5—4,6 млрд лет (по некоторым данным, 7 млрд лет). Температура ее поверхности была очень высокой — 4000 —8000 °С (по другим данным, 1000°С, т.е. температура раскаленной лавы), и по мере остывания планеты и действия гравитационных сил происходило образование земной коры из соединений различных элементов.

Первый этап возникновения жизни характеризовался процессами дегазации, которые привели к созданию атмосферы, обогащенной, возможно, азотом, аммиаком, парами воды, углекислым и угарным газами. При этом в атмосфере имелись атомы водорода, углерода, кислорода и азота, составляющие 99 % атомов, входящих в мягкие ткани любого живого организма. Чтобы атомы превратились в сложные молекулы, нужна была дополнительная энергия, которая имелась на Земле как результат вулканической деятельности, электрических грозовых разрядов, радиоактивности, ультрафиолетового излучения Солнца.
Отсутствие свободного кислорода было, вероятно, необходимым условием для возникновения жизни. Если бы свободныйкислород присутствовал на Земле в добиотический период, то, с одной стороны, он окислял бы синтезирующиеся органические вещества, а с другой — образуя озоновый слой в верхних горизонтах атмосферы, поглощал бы высокоэнергетическое ультрафиолетовое излучение Солнца. В рассматриваемый период возникновения жизни, длившийся примерно 1000 млн лет, ультрафиолет был, вероятно, основным источником энергии для синтеза органических веществ. Из водорода, азота и соединений углерода при наличии свободной энергии на Земле должны были возникать сначала простые молекулы (аммиак, метан и подобные простые соединения), которые в дальнейшем в первичном океане могли вступать в реакции между собой и с другими веществами, образуя новые соединения.

В 1953 г. американский биохимик Стенли Миллер и физик Гарольд Юри смогли экспериментально смоделировать те условия, которые существовали на Земле приблизительно 4 млрд лет назад. В специальной установке (аппарат Миллера) они подвергли смесь метана, аммиака, воды и водорода действию электрических разрядов. В результате блестящих опытов были получены аминокислоты: глицин, аланин, глутаминовая и аспаргиновая кислоты. Таким образом, предположение академика А. И.Опарина подтверждалось.

Стенли Миллер (1930-2007)

Гарольд Юри (1893-1981)

Аппарат Миллера
Второй этап состоял в дальнейших превращениях органических веществ и образовании абиогенным путем более сложных органических соединений, в том числе биологических полимеров. Американский химик С. Фокс составлял смеси аминокислот, подвергал их нагреву и получал протеиноподобные вещества. В небольших углублениях в застывающей лаве возникали водоемы, содержащие растворенные в иоде аминокислоты. Когда вода испарялась или выплескивалась на горячие камни, аминокислоты вступали в реакцию, образуя протеноиды. Если некоторые из этих протеноидов обладали каталитической активностью, то мог начаться синтез полимеров, т.е. белковоподобных молекул.
Третий этап характеризовался выделением в первичном «питательном бульоне» особых коацерватных капель, представляющих собой группы полимерных соединений. Коацерватные капли обладают некоторыми свойствами, характерными и для живой протоплазмы, например способностью избирательно адсорбировать вещества из окружающего раствора и за счет этого «расти», увеличивать свои размеры. Поскольку концентрация веществ в коацерватных каплях была в десятки раз больше, чем в окружающем растворе, возможность взаимодействия между отдельными молекулами значительно возрастала. Гидрофильные части молекул, расположенные на границе между коацерватами и раствором, поворачиваются в сторону раствора, где содержание воды больше. Гидрофобные части ориентируются внутрь коацерватов, где концентрация воды меньше. В результате поверхность коацерватов приобретает определенную структуру и в связи с этим свойство пропускать в определенном направлении одни вещества и не пропускать другие. Благодаря этому свойству концентрация некоторых веществ внутри коацерватов еще больше возрастает, других уменьшается и реакции между компонентами коацерватов приобретают определенную направленность. Коацерватные капли становятся системами, обособленными от среды. Возникают протоклетки, или протобионты. Важным этапом химической эволюции явилось образование мембранной структуры. Параллельно с появлением мембраны шло упорядочение и усовершенствование метаболизма. Одним из основных признаков живого является способность к репликации, т.е. созданию копий, не отличаемых от материнских молекул. Таким свойством обладают нуклеиновые кислоты, которые в отличие от белков способны к репликации. В коацерватах мог образовываться протеноид, способный катализировать полимеризацию нуклеотидов с образованием коротких цепочек РНК. Эти цепочки могли выполнять роль как примитивного гена, так и информационнойРНК. Уже на стадии формирования протобионтов происходил, вероятно, естественный отбор. Появление структур, способных к самовоспроизведению, репликации, изменчивости, определяет, по-видимому, четвертый этап становления жизни.
Итак, в позднем архее (приблизительно 3,5 млрд лет назад) на дне небольших водоемов или мелководных, теплых и богатых питательными веществами морей возникли первые примитивные живые организмы, которые по типу питания были гетеротрофами.
Способом обмена веществ им служило, вероятно, брожение. Часть энергии, выделяемой в этих процессах, запасается в виде АТФ. Возможно, некоторые организмы для жизненных процессов использовали и энергию окислительно-восстановительных реакций, т.е. были хемосинтетиками. Со временем происходило уменьшение запасов свободной органики в окружающей среде и преимущество получили организмы, способные синтезировать органические соединения из неорганических. Таким путем, вероятно, около 2 млрдлет назад возникли первые фототрофные организмы типа цианобактерий. Переход к автотрофному питанию имел большое значение для эволюции жизни на Земле. При этом атмосфера стала приобретать окислительный характер. Появление озонового экрана защитило первичные организмы от губительного воздействия ультрафиолетовых лучей и положило конец абиогенному (небиологическому) синтезу органических веществ.
2. Усложнение живых организмов в процессе эволюции. Многообразие живого мира на Земле и современная его организация:
Усложнение живых организмов в процессе эволюции:
Историю органического мира на Земле изучают по сохранившимся остаткам, отпечаткам и другим следам жизнедеятельности живых организмов. Она является предметом науки палеонтологии. Исходя из того, что остатки разных организмов расположены в различных пластах горных пород, была создана геохронологическая шкала, согласно которой историю Земли разделили на определенные промежутки времени: зоны, эры, периоды и века.
Эоном называют большой промежуток времени в геологической истории, объединяющий несколько эр. В настоящее время выделяют только два зона: криптозой (скрытая жизнь) и фанерозой (явная жизнь).
Эра - это промежуток времени в геологической истории, являющийся подразделением эона, объединяющий, в свою очередь, периоды. В криптозое выделяют две эры (архей и протерозой), тогда как в фанерозое - три (палеозой, мезозой и кайнозой).
Важную роль в создании геохронологической шкалы сыграли руководящие ископаемые — остатки организмов, которые были многочисленны в определенные промежутки времени и хорошо сохранились.
Развитие жизни в криптозое. Архей и протерозой составляют большую часть истории жизни (период 4,6 млрд лет - 0,6 млрд лет назад), однако сведений о жизни в тот период недостаточно. Первые остатки органических веществ биогенного происхождения имеют возраст около 3,8 млрд лет, а прокариотические организмы существовали уже 3,5 млрд лет назад. Первые прокариоты входили в состав специфических экосистем - цианобактериальных матов, благодаря деятельности которых образовались специфические осадочные породы строматолиты («каменные ковры»).
Понять жизнь давних прокариотических экосистем помогло открытие их современных аналогов - строматолитов в заливе Шарк-Бей в Австралии и специфических пленок на поверхности почвы в заливе Сиваш в Украине. На поверхности цианобактериальных матов расположены фото- синтезирующие цианобактерии, а под их слоем - чрезвычайно разнообразные бактерии других групп и архей. Минеральные вещества, которые оседают на поверхность мата и образовываются за счет его жизнедеятельности, откладываются пластами (приблизительно 0,3 мм в год). Такие примитивные экосистемы могут существовать лишь в непригодных для жизни других организмов местах, и действительно, оба вышеупомянутые местообитания характеризуются чрезвычайно высокой соленостью.
Многочисленные данные свидетельствуют о том, что поначалу Земля имела атмосферу возобновляемого характера, в состав которой входили: углекислый газ, водяной пар, оксид серы, а также угарный газ, водород, сероводород, аммиак, метан и т. п. Первые организмы Земли были анаэробами, однако благодаря фотосинтезу цианобактерий в среду выделялся свободный кислород, который сначала быстро связывался с восстановителями, находящимися в среде, и лишь после связывания всех восстановителей среда начала приобретать окислительные свойства. О таком переходе свидетельствуют отложение окисленных форм железа — гематита и магнетита.
Около 2 млрд лет назад в результате геофизических процессов практически все несвязанное в осадочных породах железо переместилось к ядру планеты, а кислород начал накапливаться в атмосфере из-за отсутствия этого элемента - произошла «кислородная революция». Она явилась переломным этапом в истории Земли, который повлек за собой не только смену состава атмосферы и образование озонового экрана в атмосфере - главной предпосылки для заселения суши, но и состава пород, формирующихся на поверхности Земли.
В протерозое произошло и другое важное событие - возникновение эукариот. В последние годы удалось собрать убедительные доказательства теории эндосимбиогенетического происхождения эукариотической клетки - путем симбиоза нескольких прокариотических клеток. Вероятно, «главным» предком эукариот стали архей, которые перешли к поглощению пищевых частиц путем фагоцитоза. Наследственный аппарат переместился вглубь клетки, сохранив, тем не менее, связь с мембраной благодаря переходу внешней мембраны возникшей ядерной оболочки в мембраны эндоплазматической сети.
Поглощенные клеткой бактерии могли не перевариваться, а оставаться живыми и продолжать свое функционирование. Считают, что митохондрии ведут свое происхождение от пурпурных бактерий, утративших способность к фотосинтезу и перешедших к окислению органических веществ. Симбиоз с другими фотосинтезирующими клетками привел к возникновению пластид у растительных клеток. Вероятно, жгутики эукариотических клеток возникли вследствие симбиоза с бактериями, которые, подобно современным спирохетам, были способны к извивающимся движениям. Поначалу наследственный аппарат эукариотических клеток был устроен приблизительно так же, как у прокариот, и лишь позднее, вследствие необходимости управления большой и сложной клеткой, образовались хромосомы. Геномы внутриклеточных симбионтов (митохондрий, пластид и жгутиков) в целом сохранили прокариотическую организацию, но большая часть их функций перешла к ядерному геному.
Эукариотические клетки возникали неоднократно и независимо друг от друга. Например, красные водоросли возникли в результате симбиогенеза с цианобактериями, а зеленые водоросли - с бактериями-прохлорофитами.
Остальные одномембранные органеллы и ядро эукариотической клетки, согласно эндомем- бранной теории, возникли из впячиваний мембраны прокариотической клетки.
Точное время появления эукариот неизвестно, поскольку уже в отложениях возрастом около 3 млрд лет присутствуют отпечатки клеток, имеющих похожие размеры. Точно эукариоты зафиксированы в породах возрастом около 1,5-2 млрд лет, но только после кислородной революции (около 1 млрд лет назад) сложились условия, благоприятные для них.
В конце протерозойской эры (не менее 1,5 млрд лет назад) уже существовали и многоклеточные эукариотические организмы. Многоклеточность, как и эукариотическая клетка, неоднократно возникала у разных групп организмов.
Существуют различные взгляды на происхождение многоклеточных животных. По одним данным их родоначальниками были многоядерные, подобные инфузориям, клетки, которые затем распались на отдельные одноядерные клетки.
Другие гипотезы связывают происхождение многоклеточных животных с дифференцировкой клеток колониальных одноклеточных. Расхождения между ними касаются возникновения слоев клеток у первоначального многоклеточного животного. Согласно гипотезы гастреи Э. Геккеля, то происходит путем впячивания одной из стенок однослойного многоклеточного организма, как у кишечнополостных. В противовес ей И. И. Мечников сформулировал гипотезу фагоцителлы, считая предками многоклеточных однослойные шарообразные колонии наподобие вольвокса, которые поглощали пищевые частицы путем фагоцитоза. Клетка, захватившая частицу, теряла жгутик и переходила вглубь организма, где и осуществляла пищеварение, а по окончании процесса возвращалась на поверхность. Со временем произошло разделение клеток на два слоя с определенными функциями - внешний обеспечивал движение, а внутренний - фагоцитоз. Такой организм И. И. Мечников назвал фагоцителлой.
В течение продолжительного времени многоклеточные эукариоты проигрывали в конкурентной борьбе прокариотическим организмам, однако в конце протерозоя (800-600 млн лет тому) вследствие резкого изменения условий на Земле - снижения уровня морей, роста концентрации кислорода, уменьшения концентрации карбонатов в морской воде, регулярных циклов похолодания - многоклеточные эукариоты получили преимущества над прокариотами. Если до этого времени встречались только отдельные многоклеточные растения и, возможно, грибы, то с этого момента в истории Земли известны и животные. Из возникших в конце протерозоя фаун лучше других изучены эдиакарская и вендская. Животных вендского периода принято включать в состав особой группы организмов или относить к таким типам, как кишечнополостные, плоские черви, членистоногие и др. Однако ни у одной из этих групп нет скелетов, что может свидетельствовать об отсутствии хищников.
Развитие жизни в палеозойской эре. Палеозойская эра, длившаяся более 300 млн лет, делится на шесть периодов: кембрийский, ордовикский, силурийский, девонский, каменноугольный (карбон) и пермский.
В кембрийском периоде суша состояла из нескольких материков, расположенных преимущественно в Южном полушарии. Самыми многочисленными фотосинтезирующими организмами в этот период были цианобактерии и красные водоросли. В толще воды жили фораминиферы и радиолярии. В кембрии появляется огромное количество скелетных животных организмов, о чем свидетельствуют многочисленные ископаемые остатки. Эти организмы относились примерно к 100 типам многоклеточных животных, как современным (губки, кишечнополостные, черви, членистоногие, моллюски), так и исчезнувшим, например: огромный хищник аномалока- рис и колониальные граптолиты, которые плавали в толще воды или были прикреплены ко дну. Суша на протяжении кембрия оставалась почти незаселенной, однако процесс почвообразования уже начали бактерии, грибы и, возможно, лишайники, а в конце периода на сушу вышли мало- щетинковые черви и многоножки.
В ордовикском периоде уровень вод Мирового океана поднялся, что привело к затоплению материковых низменностей. Основными продуцентами в этот период были зеленые, бурые и красные водоросли. В отличие от кембрия, в котором рифы строили губки, в ордовике их сменяют коралловые полипы. Расцвет переживали брюхоногие и головоногие моллюски, а также трилобиты (ныне вымершие родственники паукообразных). В этом периоде впервые зафиксированы и хордовые, в частности бесчелюстные. В конце ордовика произошло грандиозное вымирание, которое уничтожило около 35 % семейств и более 50 % родов морских животных.
Силурийский период характеризуется усилением горообразования, которое привело к осушению материковых платформ. Ведущую роль в фауне беспозвоночных силура играли головоногие моллюски, иглокожие и гигантские ракоскорпионы, тогда как среди позвоночных сохраняется большое разнообразие бесчелюстных и появляются рыбы. В конце периода на сушу вышли первые сосудистые растения - риниофиты и плауновидные, которые начали колонизацию мелководья и приливно-отливной зоны побережий. На сушу вышли и первые представители класса паукообразных.
В девонском периоде вследствие поднятия суши образовались большие мелководья, которые пересыхали и даже промерзали, поскольку климат становился еще более континентальным, чем в силуре. В морях преобладают кораллы и иглокожие, тогда как головоногие моллюски представлены спирально закрученными аммонитами. Среди позвоночных девона расцвета достигли рыбы, причем на смену панцирным пришли и хрящевые, и костные, а также двоякодышащие и кисте- перые. В конце периода появляются первые амфибии, которые сначала жили в воде.
В среднем девоне на суше появились первые леса из папоротников, плаунов и хвощей, которые были заселены червями и многочисленными членистоногими (многоножками, пауками, скорпионами, бескрылыми насекомыми). В конце девона появились первые голосеменные. Освоение суши растениями привело к уменьшению выветривания и усилению почвообразования. Закрепление почв привело к возникновению русел рек.
В каменноугольном периоде суша была представлена двумя материками, разделенными океаном, а климат стал заметно более теплым и влажным. К концу периода произошло небольшое поднятие суши, а климат сменился более континентальным. В морях господствовали фораминиферы, кораллы, иглокожие, хрящевые и костные рыбы, а пресные водоемы населяли двухстворчатые моллюски, ракообразные и разнообразные земноводные. В середине карбона возникли мелкие насекомоядные рептилии, а среди насекомых появились крылатые (тараканы, стрекозы).
Для тропиков были характерны заболоченные леса, в которых доминировали гигантские хвощи, плауны и папоротники, отмершие остатки которых образовали впоследствии залежи каменного угля. В середине периода в умеренной зоне, благодаря их независимости от воды в процессе оплодотворения и наличию семени, началось распространение голосеменных.
Пермский период отличался слиянием всех материков в единый суперконтинент Пангею, отступлением морей и усилением континентальности климата до такой степени, что во внутренних районах Пангеи образовались пустыни. К концу периода на суше почти исчезли древовидные папоротники, хвощи и плауны, а господствующее положение заняли засухоустойчивые голосеменные.
Несмотря на то, что крупные амфибии еще продолжали существовать, возникли разные группы рептилий, в том числе крупных растительноядных и хищных. В конце перми произошло самое большое вымирание в истории жизни, так как исчезли многие группы кораллов, трилобиты, большинство головоногих, рыб (в первую очередь хрящевых и кистеперых), а также амфибий. Морская фауна потеряла при этом 40-50% семейств и около 70% родов.
Развитие жизни в мезозое. Мезозойская эра продолжалась около 165 млн лет и характеризовалась поднятием суши, интенсивным горообразованием и снижением влажности климата. Она делится на три периода: триасовый, юрский и меловой.
В начале триасового периода климат был засушливым, однако позднее вследствие поднятия уровня морей он стал более влажным. Среди растений преобладали голосеменные, папоротники и хвощи, однако древесные формы споровых практически полностью вымерли. Высокого развития достигли некоторые кораллы, аммониты, новые группы фораминифер, двухстворчатых моллюсков и иглокожих, тогда как разнообразие хрящевых рыб уменьшилось, изменились и группы костных рыб. Господствовавшие на суше рептилии начали осваивать и водную среду, как ихтиозавры и плезиозавры. Из пресмыкающихся триаса до нашего времени дожили крокодилы, гаттерии и черепахи. В конце триаса появились динозавры, млекопитающие и птицы.
В юрском периоде суперконтинент Пангея раскололся на несколько меньших. Большая часть юры была очень влажной, а к его концу климат стал более засушливым. Доминирующей группой растений были голосеменные, из которых от того времени сохранились секвойи. В морях процветали моллюски (аммониты и белемниты, двухстворчатые и брюхоногие), губки, морские ежи, хрящевые и костные рыбы. Крупные амфибии практически полностью вымерли в юрском периоде, однако появились современные группы земноводных (хвостатые и бесхвостые) и чешуйчатых (ящериц и змей), возросло разнообразие млекопитающих. К концу периода возникли и возможные предки первых птиц — археоптериксы. Однако во всех экосистемах доминировали пресмыкающиеся — ихтиозавры и плезиозавры, динозавры и летающие ящеры — птерозавры.
Меловой период получил название в связи с образованием мела в осадочных породах того времени. На всей Земле, кроме приполярных областей, был стойкий теплый и влажный климат. В этом периоде возникли и приобрели широкое распространение покрытосеменные, вытеснявшие голосеменных, что повлекло за собой резкое увеличение разнообразия насекомых. В морях, помимо моллюсков, костистых рыб, плезиозавров, вновь появилось огромное количество форами-нифер, раковинки которых и образовали залежи мела, а на суше преобладали динозавры. Лучше приспособленные к воздушной среде птицы начали постепенно вытеснять летающих ящеров.
В конце периода произошло глобальное вымирание, в результате которого исчезли аммониты, белемниты, динозавры, птерозавры и морские ящеры, древние группы птиц, а также некоторые голосеменные. С лица Земли в целом исчезло около 16% семейств и 50% родов животных. Кризис в конце мела связывают с падением большого метеорита в Мексиканский залив, однако он, скорее всего, не был единственной причиной глобальных изменений. В ходе последующего похолодания выжили только небольшие рептилии и теплокровные млекопитающие.
Развитие жизни в кайнозое. Кайнозойская эра началась около 66 млн лет назад и продолжается до настоящего времени. Она характеризуется господством насекомых, птиц, млекопитающих и покрытосеменных растений. Кайнозой делят на три периода - палеоген, неоген и антропоген - последний из которых является самым коротким в истории Земли.
В раннем и среднем палеогене климат оставался теплым и влажным, к концу периода стало прохладнее и суше. Доминирующей группой растений стали покрытосеменные, однако, если в начале периода преобладали вечнозеленые леса, то в конце появилось много листопадных, а в засушливых зонах образовались степи.
Среди рыб господствующее положение заняли костистые рыбы, а количество видов хрящевых, несмотря на их заметную роль в соленых водоемах, незначительно. На суше из рептилий сохранились только чешуйчатые, крокодилы и черепахи, тогда как млекопитающие заняли большую часть их экологических ниш. В середине периода появились основные отряды млекопитающих, в том числе насекомоядные, хищные, ластоногие, китообразные, копытные и приматы. Изоляция материков сделала фауну и флору географически более разнообразными: Южная Америка и Австралия стали центрами развития сумчатых, а другие материки - плацентарных млекопитающих.
Неогеновый период. Земная поверхность в неогене приобрела современный вид. Климат стал более прохладным и сухим. В неогене уже сформировались все отряды современных млекопитающих, а в африканских саванах возникло семейство Гоминид и род Человек. К концу периода в приполярных областях континентов распространились хвойные леса, появились тундры, а степи умеренного пояса заняли злаки.
Четвертичный период (антропоген) характеризуется периодическими сменами оледенений и потеплений. Во время оледенений высокие широты покрывались ледниками, резко снижался уровень океана, суживались тропический и субтропический пояса. На близлежащих к ледникам территориях устанавливался холодный и сухой климат, который способствовал формированию холодоустойчивых групп животных - мамонтов, гигантских оленей, пещерных львов и др. Сопутствовавшее процессу оледенения снижение уровня Мирового океана привело к образованию сухопутных мостов между Азией и Северной Америкой, Европой и Британскими островами и т. д. Миграции животных, с одной стороны, привели к взаимообогащению флор и фаун, а с другой, к вытеснению реликтов пришельцами, например, сумчатых и копытных в Южной Америке. Эти процессы, однако, не затронули Австралию, оставшуюся изолированной.
В целом, периодические изменения климата привели к формированию чрезвычайно обильного видового разнообразия, характерного для нынешнего этапа эволюции биосферы, а также оказали влияние на эволюцию человека. На протяжении антропогена несколько видов рода Человек расселились из Африки в Евразию. Около 200 тысяч лет назад в Африке возник вид Человек разумный, который после продолжительного периода существования в Африке около 70 тысяч лет назад вышел в Евразию и около 35-40 тыс. лет назад - в Америку. После периода сосуществования с близкородственными видами он вытеснил их и расселился по всей территории земного шара.
Около 10 тыс. лет назад хозяйственная деятельность человека в умеренно теплых областях земного шара начала оказывать влияние как на облик планеты (распашка земель, выжигание лесов, перевыпас пастбищ, опустынивание и т. д.), так и на животный и растительный мир вследствие сокращения ареалов их обитания и истребления, и вступил в действие антропогенный фактор.
Происхождение человека. Человек как вид, его место в системе органического мира. Гипотезы происхождения человека. Движущие силы и этапы эволюции человека. Человеческие расы, их генетическое родство. Биосоциальная природа человека. Социальная и природная среда, адаптации к ней человека.
----------------------
Многообразие живого мира на Земле и современная его организация:
Все многообразие живого мира практически невозможно выразить в количественном эквиваленте. По этой причине систематики объединили их в группы на основании определенных признаков.
Каждый вид, существующий на планете, индивидуален и неповторим. Однако многие из них имеют целый ряд сходных черт строения. Именно по этим признакам все живое можно объединить в таксоны. В современный период ученые выделяют пять Царств.
Многообразие живого мира (фото демонстрирует некоторых его представителей) включает Растения, Животные, Грибы, Бактерии и Вирусы. Последние из них не имеют клеточного строения и по этому признаку относятся к отдельному Царству.
Молекула вирусов состоит из нуклеиновой кислоты, которая может быть представлена как ДНК, так и РНК. Вокруг них располагается белковая оболочка. С таким строением данные организмы способны осуществлять только единственный признак живых существ - размножаться самосборкой внутри организма хозяина.
Все бактерии являются прокариотами. Это значит, что в их клетках нет оформленного ядра. Их генетический материал представлен нуклеоидом - кольцевыми молекулами ДНК, скопления которых находятся прямо в цитоплазме.
Растения и животные отличаются способом питания. Первые способны сами синтезировать органические вещества в ходе фотосинтеза. Такой способ питания называется автотрофным. Животные поглощают уже готовые вещества. Такие организмы называют гетеротрофами.
Грибы обладают признаками как растений, так и животных. К примеру, они ведут прикрепленный образ жизни и неограниченный рост, но не способны к фотосинтезу. Свойства живой материи.
А по каким признакам, вообще, организмы называют живыми? Ученые выделяют целый ряд критериев. Прежде всего, это единство химического состава. Вся живая материя образована органическими веществами. К ним относятся белки, липиды, углеводы и нуклеиновые кислоты. Все они являются естественными биополимерами, состоящими из определенного количества повторяющихся элементов.
К признакам живых существ также принадлежат питание, дыхание, рост, развитие, наследственная изменчивость, обмен веществ, размножение, способность к адаптации. Каждый таксон характеризуется своими особенностями.
К примеру, растения произрастают неограниченно, в течение всей жизни. А вот животные увеличиваются в размерах только до определенного времени. То же самое касается и дыхания. Принято считать, что этот процесс происходит только при участии кислорода. Такое дыхание называется аэробным. Но вот некоторые бактерии могут окислять органические вещества и без наличия кислорода - анаэробно.
Многообразие живого мира: уровни организации и основные свойства Указанными признаками живого обладает и микроскопическая бактериальная клетка, и огромный голубой кит. Кроме того, все организмы в природе взаимосвязаны непрерывным обменом веществ и энергии, а также являются необходимыми звеньями в цепях питания. Несмотря на многообразие живого мира, уровни организации предполагают наличие только определенных физиологических процессов. Они ограничиваются особенностями строения и видовым разнообразием. Рассмотрим каждый из них подробнее.
Молекулярный уровень Многообразие живого мира наряду с его уникальностью определяется именно этим уровнем. Основу всех организмов составляют белки, структурным элементов которых являются аминокислоты. Количество их невелико - около 170. Но в состав белковой молекулы входит всего 20. Их сочетание обуславливает бесконечное разнообразие белковых молекул - от запасного альбумина птичьих яиц до коллагена мышечных волокон. На этом уровне осуществляется рост и развитие организмов в целом, хранение и передача наследственного материала, обмен веществ и превращение энергии.
Клеточный и тканевый уровень Молекулы органических веществ формируют клетки. Многообразие живого мира, основные свойства живых организмов на этом уровне уже проявляются в полном объеме. В природе широко распространены одноклеточные организмы. Это могут быть как бактерии, так и растения, и животные. У таких существ клеточный уровень соответствует организменному. На первый взгляд может показаться, что их строение достаточно примитивно. Но это совсем не так. Только представьте: одна клетка выполняет функции целого организма! К примеру, инфузория туфелька осуществляет движение с помощью жгутика, дыхание через всю поверхность, пищеварение и регуляцию осмотического давления посредством специализированных вакуолей. Известен у этих организмов и половой процесс, который происходит в форме конъюгации. У многоклеточных организмов формируются ткани. Эта структура состоит из клеток, сходных по строению и функциям.
Организменный уровень В биологии многообразие живого мира изучается именно на этом уровне. Каждый организм является единым целым и работает согласовано. Большинство из них состоит их клеток, тканей и органов. Исключением являются низшие растения, грибы и лишайники. Их тело образовано совокупностью клеток, которые не формируют тканей и называется слоевищем. Функцию корней в организмах такого типа выполняют ризоиды.
Популяционно-видовой и экосистемный уровень Наименьшей единицей в систематике является вид. Это совокупность особей, обладающих рядом общих черт. Прежде всего, это морфологические, биохимические особенности и способность к свободному скрещиванию, позволяющие обитать данным организмам в пределах одного ареала и давать плодовитое потомство.
Современная систематика насчитывает более 1,7 млн. видов. Но в природе они не могут существовать разрозненно. В пределах определенной территории обитает сразу несколько видов. Это и определяет многообразие живого мира. В биологии совокупность особей одного вида, которые обитают в пределах определенного ареала, называются популяцией. От подобных групп они изолированы определенными природными барьерами. Это могут быть водоемы, горные или лесные массивы. Каждая популяция характеризуется своим разнообразием, а также половой, возрастной, экологической, пространственной и генетической структурой. Но даже в пределах отдельно взятого ареала, видовое разнообразие организмов достаточно велико. Все они приспособлены к обитанию в определенных условиях и тесно связаны трофически. Это означает, что каждый вид является источником питания для другого. В результате формируется экосистема, или биоценоз. Это уже совокупность особей уже разных видов, связанных местом обитания, круговоротом веществ и энергии.
Биогеоценоз Но со всеми организмами постоянно взаимодействуют факторы неживой природы. К ним относятся температурный режим воздуха, соленость и химический состав воды, количество влаги и солнечного света. Все живые существа находятся в зависимости от них и не могут существовать без определенных условий. К примеру, растения питаются только при наличии солнечной энергии, воды и углекислого газа. Это условия фотосинтеза, в ходе которого синтезируются необходимые им органические вещества. Совокупность биотических факторов и неживой природы называются биогеоценозом. Что такое биосфера Многообразие живого мира в самом широком масштабе представлено биосферой. Это глобальная природная оболочка нашей планеты, объединяющая все живое. Биосфера имеет свои границы. Верхняя, расположенная в атмосфере, ограничена озоновым слоем планеты. Он расположен на высоте 20 - 25 км. Данный слой поглощает вредное ультрафиолетовое излучение. Выше него жизнь просто невозможна. На глубине до 3 км находится нижняя граница биосферы. Здесь она ограничена наличием влаги. Так глубоко способны обитать только анаэробные бактерии. В водной оболочке планеты - гидросфере, жизнь найдена на глубине 10-11 км. Итак, живые организмы, населяющие нашу планету в разных природных оболочках, обладают рядом характерных свойств. К ним относят их способность к дыханию, питанию, движению, размножению и т. д. Многообразие живых организмов представлено разными уровнями организации, каждый из которых отличается уровнем сложности структуры и физиологических процессов.

