 2020-07-12
2020-07-12 115
115Материалы и методы
Микрометеорология. Полевые работы проводились с 10 по 21 июня 1999 г. в Национальном парке Кишкунсаг, примерно в 70 км к югу от Будапешта, недалеко от Фюлефазы, Венгрия (19–140 ° E; 47–300 ° с.ш.), на высоте 130 м над уровнем моря. Осадки и преобладающие температуры в Кишкунсаге достаточны для поддержания древесных растений, но высокая гидравлическая проводимость песчаных почв значительно усиливает сезонное высыхание, благоприятствуя доминированию пучковых трав. В этом районе Кишкунсага распространен обширный покров эктоидного мха Tortula ruralis, биомасса которого, как сообщается, превышает биомассу сообщества сосудистых растений. (Tuba et al. 1996). Альбедо и поверхностная сетчатая радиация были измерены на одной области, преимущественно покрытой T.uralis, и на одной области с голым песком, причем оба с покровом сосудистых растений <10%. Обе станции отбора проб имели одинаковые уклоны и были расположены в пределах 20 м друг от друга, чтобы гарантировать, что изменения в разнице между альбедо в двух областях будут в основном связаны с изменениями в отображении площади листа мха. На каждой станции кремниевый фотодиодный пиранометр (LiCOR Model 200SA, Линкольн, Небраска) был ориентирован непосредственно вниз на 70 см над поверхностью земли для оценки альбедо. Пиранометр был прикреплен к нижней части штатива, держащего плечо мачты выровненного сетевого радиометра (Radiation Energy Balance Systems, Сиэтл, Вашингтон, США), для количественной оценки чистого потока излучения (кВтч/м²). Сетчатые радиометры были ориентированы параллельно 180° от магнитного севера на высоте 1 м над уровнем земли. Общее солнечное излучение (кВтч/м²) измеряли с помощью выровненного пиранометра LiCOR Si-200SA, плотность потока фотосинтетических фотонов (PPFD; мкмоль/с/м2) с помощью квантового датчика LiCOR 190 SA и скорость ветра (м/с) с помощью чашечного анемометра (модель 014A, MetOne Instruments, Grants Pass, Орегон, США). Эти инструменты были размещены на высоте 1 м над уровнем земли на удалении 15 м от чистых радиационных станций. Все инструменты были повторно выровнены по мере необходимости три раза в день в течение всего исследования. Радиационные данные регистрировали каждую минуту, затем усредняли и сохраняли каждые пять минут, 24 часа в сутки в течение всего периода исследования, используя регистраторы данных LiCOR 1400 (LiCOR Instruments, Линкольн, Небраска, США). Альбедо было рассчитано как А=1−((полное облучение−облучение от поверхности)/полное облучение). Данные о скорости ветра использовались для коррекции любых конвективных теплообменных эффектов на сетчатых радиометрах с использованием корректирующих соотношений, предоставленных изготовителем. Air temperature, relative humidity, and dew point temperature were measured at the moss-dominated station with a Hobo Pro8 datalogger (Onset Computer Corp, Brookline, Massachusetts, USA). Температура воздуха измерялась на высоте 1 м над поверхностью земли. Относительная влажность и температура точки росы измерялись на высоте 5 см над землей, чтобы более точно отразить динамику атмосферной влажности микроклимата мохового ковра. Температуру мохового коврика измеряли, осторожно вставляя встроенный в эпоксидную смолу термистор / регистратор данных (TidBit, Onset Computer Corp., Бруклин, Массачусетс) под Моховой коврик, заботясь о том, чтобы обеспечить контакт между датчиком и нижней стороной коврика. Эти микрометеорологические измерения производились каждую минуту и в среднем длились более 5 минут.
Физиологические измерения. Импульсно-амплитудно-модулированная (ПАМ) флуоресценция хлорофилла использовалась для оценки суточной физиологической активности полностью регидратированного T. ruralis в течение трех дней (14, 17 и 19 июня – Рис.1). Перед каждым отбором проб новый участок мха площадью около 100 см2 опрыскивали водой в течение двух минут, а затем давали увлажниться еще в течение трех минут. Эти области были близки к полусферическому виду приборов, используемых для контроля микрометеорологического и поверхностного отражения невозмущенных областей, но не находились внутри него, как описано выше. Этот протокол смачивания применялся во избежание проблем с сушкой ткани, которые сильно снижают силу сигнала флуоресценции и отношения сигнал/шум и могут давать сильно завышенные и ненадежные значения коэффициентов флуоресценции хлорофилла. (E.P. Hamerlynck, неопубликованные данные). Протокол позволил нам показать физиологическое состояние растений сразу после регидратации с естественных уровней сухости при температуре окружающей среды (Tuba et al. 1996). Пять-шесть отростков T.ruralis аккуратно помещали между двумя полностью насыщенными кусочками папиросной бумаги в камере для адаптации к темноте и адаптировали к темноте в течение минимум 15 минут. На каждом временном интервале было сделано по пять независимых выборок. Каждый раз регидратировалась новая область, чтобы обеспечить независимость измерений для статистического анализа. Исходная флуоресценция (Fo) усреднялась в течение двух секунд после десятисекундного воздействия модулирующего луча. Максимальная флуоресценция (Fm) была достигнута при освещении 3000 мкмоль/с*м² белым светом в течение 0,8 сек. с помощью лампы высокой интенсивности, направленной через волоконно-оптический кабель. Максимальная фотохимическая эффективность PSII была определена как Fv / Fm, где Fv = Fm-Fo (Schreiber et al. 1998). Fv / Fm оценивался с двухчасовыми интервалами в течение дня, за исключением 17 июня, когда технические проблемы ограничили нас измерениями рано утром, в полдень и на закате. В каждом из этих трех раз во все дни сразу после темноадаптированных измерений оценивались светоотражающие кривые фотохимического тушения (qP, индикатор восстановительного состояния первичного акцептора электронов Qa), нерадиационная тепловая диссипация энергии возбуждения PSII (NPQ) и эффективный квантовый выход PSII (φPSII) по формулам Schreiber et al. (1998). Все измерения производились в герметичной камере, прикрепленной к створчатому зажиму, причем единственным источником света был прибор. Актинический свет на уровне растений был увеличен до 25, 60, 125, 250, 350, и 725 мкмоль/с*м² PPFD, что было проверено с помощью квантового датчика LiCOR 180. Растениям давали уравновеситься в течение двух минут на каждом уровне освещенности, а затем освещали в течение 0,8 сек насыщающим лучом, чтобы обеспечить адаптированную к свету максимальную флуоресценцию (F0m). Через тридцать секунд после оценки F0 m источник актинического света был выключен, и слабый луч дальнего красного света использовался для оценки F0 o для использования при корректировке оценок параметров гашения.
Статистический анализ. Для выявления влияния даты и времени суток (раннее утро, полдень и закат солнца) на Fv/Fm использовался двусторонний дисперсионный анализ (ANOVA; GLM протокол SYSTAT v8.0; SPSS Software, Чикаго, Иллинойс, США). Двухсторонние повторные измерения ANOVA (Systat v8.0) были использованы для проверки эффектов взаимодействия даты, времени суток и даты × времени для параметров тушения, полученных из кривых световой реакции. Особый интерес в этом анализе представляли внутрисубъектные эффекты, поскольку они отражали бы межсуточные и внутрисуточные различия реакций на актинический уровень освещенности, повторную меру. В обоих анализах дату рассматривали как случайный эффект, а время - как фиксированный эффект, используя взаимодействия дата-репликация и дата-время-репликация, соответственно, как термины ошибки. Все данные были преобразованы в арксинус для соответствия предположениям ANOVA. (Zar 1974). р ≤ 0,05 считалось значимым. Линейная и нелинейная регрессия (Sigmaplot v4. 01, SPSS) были использованы для установления взаимосвязей между параметрами окружающей среды и альбедо, а также между Fv/Fm и альбедо. Для минимизации погрешности датчиков от низких углов солнца использовались данные микрометеорологии и альбедо от 0600 до 1800. Для учета изменений альбедо во время сбора образцов растений данные по альбедо были объединены в течение 15 мин из пятиминутных периодов до, во время и после адаптации к темноте. R2 больше или равно 0,70, что соответствует p ≤ 0,0001, считалось значимым для этих анализов.
Результаты
Три дня выборки значительно различались в условиях радиации и влажности (рис. 1). 14 июня чистая радиация редко и лишь временно превышала 400 Вт/м2 в течение утра, затем достигла высоких положительных значений (600 Вт/м2) в условиях прояснения к середине дня и постепенно снизилась до значений около 100 Вт/м2 к закату солнца. Температура мохового коврика была в пределах от 1 до 3 C от точки росы в течение всего утра, а затем значительно выше температуры точки росы (значения от 5 до 15°C выше) в течение дня (рис.1). 17 июня весь день было прохладно, влажно и дождливо. Чистая радиация мохового ковра была отрицательной или очень низкой в течение раннего утра (от -15 до 25 Вт/м2); увеличилась до 175 Вт/м2, ненадолго достигнув 600 Вт/м2 к полудню; и вернулась к низким положительным или отрицательным значениям до захода солнца. Температура мохового ковра редко превышала точку росы более чем на 1 или 2° C. Утро 19 июня было относительно прохладным и ясным, но сухим. Чистая радиация мохового ковра быстро возросла с −7 Вт/м2 на восходе солнца до 340 Вт/м2 к 08:00 и достигла полуденного максимума в 645 Вт/м2. Температура мата оставалась значительно выше точки росы в течение всего дня (рис.1).
В дни с ярко выраженными периодами высыхания (14 и 19 июня) альбедо с моховым ковриком находилось в тесной параллели как с выходом температуры матов из Tроса (рис. 1), так и с относительной влажностью. Альбедо показал более слабую связь с фотосинтетической плотностью потока фотонов (рис. 2). В таких условиях Fv/Fm внимательно следили за изменениями альбедо. В устойчивых влажных условиях (17 июня) альбедо и моховый мат показали незначительные суточные колебания (рис. 2). Fv/Fm был значительно выше 17 июня (0,60±0,014 SE), чем в дни с периодами высыхания (0,5 ±0,023 SE и 0,52±0,26 SE для 14 и 19 июня соответственно; Таблица 1). Fv/Fm было выше утром (0,61±0,013 SE), чем в полдень или вечер (0,51±0,022 SE и 0,51±0,025 SE соответственно), и не показало значительных изменений во времени (таблица 1).
Моховой коврик альбедо показала сигмоидальной отношения к относительной влажности (RР = 0.13+0.031/(1 + е−((альбедо−59.7)/5.43)) и к разнице между температурой мха-мата и Tроса (Тмат−Tроса=0.13 +0.029/(1+ е−((альбедо−9.72)/-1.166)); Рис. 3). Обе взаимосвязи были значимыми (R2 = 0,72 и 0,71; MS = 0,22 и 0,21 соответственно; p ≤ 0,0001; df = 3,430). Сигмоидальные модели объясняли на 13% и 7% больше вариаций, чем соответствующие линейные модели (R2 = 0,59 и 0,64 соответственно). Альбедо не показало значимой линейной связи с PPFD (линейная регрессия R2 =0,40).
Объединенные по всем датам выборки, Fv/Fm показали сигмоидальные отношения с альбедо (рис. 4; Fv/Fm=0,457 + 0,148 / (1 + e -((альбедо-0,144) / 0,0012)). Это соотношение было значительным (скорректированное R2 = 0,96, MS = 0,0195; F=99,50; ≤0,0001; df = 3,9), что составляет на 32% больше вариаций, чем линейное соотношение (R2 = 0,64; MS = 0,0385; F = 19,44; p =0,001; df=1,11). При более высоком альбедо (от 0,15 до 0,16) Fv/Fm занимал узкий диапазон относительно высоких значений (от 0,58 до 0,65). При низком альбедо, равном 0,15, Fv/Fm резко уменьшалось, достигая минимума от 0,42 до 0,47. Промежуточные значения Fv / Fm были получены в результате незначительного дневного повышения, наблюдавшегося 14 июня (Рис.2).
Фотохимическое гашение (qP), нерадиационная диссипация (NPQ) и видимый квантовый выход PSII (φPSII) продемонстрировали значительные интерактивные эффекты даты и времени (таблица 1). Эти взаимодействия были обусловлены различным суточным поведением при насыщении или субнасыщении уровней света между датами отбора проб (Рис. 5). Фотохимическое гашение показало выраженное увеличение интенсивности насыщающего света в течение полуденного и вечернего периодов отбора проб 14 июня, что не было заметно в другие даты отбора проб (рис. 5). NPQ показал большую степень суточной вариабельности, чем qP. При уровне освещенности не менее 250 мкмоль/с*м² значения NPQ по утрам были одинаковыми в разные дни (рис. 5). В течение нескольких дней утренние значения NPQ были аналогичны дневным и вечерним значениям 14 июня, но утренние значения были заметно ниже полуденных значений 17 июня и немного выше дневных и вечерних значений 19 июня. Суточное снижение φPSII наблюдалось при более высокой интенсивности света вечером 17 июня и при более низкой интенсивности света (25-250 мкмоль/с*м²) днем и вечером 19 июня (Рис.5).
Обсуждение результатов
Наши результаты свидетельствуют о том, что быстрое закрытие побега у T. ruralis (Рис.2 и 3) приводит к эффективной степенной зависимости между Fv/Fm, показателем максимальной фотохимической эффективности, и альбедо у HDT-мха T. ruralis (рис. 4). Это подтверждает лабораторные эксперименты по регидратации, показывающие, что восстановление Fv/Fm после длительного высушивания происходит почти мгновенно, даже у относительно чувствительных к высушиванию бриофитов. (Sass et al. 1995; Csintalan et al. 1999). Короткое время регидратации, которое мы допустили в полевых условиях, меньше, чем требуется для достижения максимальной скорости фотосинтеза, и поэтому лучше всего считается, что оно отражает физиологический потенциал во время восстановления после обезвоживания. (Tuba et al. 1996; Csintalan et al. 1998). Самый высокий Fv/Fm, отмеченный здесь, довольно низок, но находится в пределах диапазонов, отмеченных для T. ruralis и других засухоустойчивых растений. (Sass et al. 1995; Csintalan et al. 1999). Возможно, что низкий Fv / Fm отражает конститутивную защитную стратегию фото/высыхания, которая высоко выражена в этих растениях. (Osmond & Grace 1994; Csintalan et al. 1999). Реакция ступенчатой функции на доступность воды ясно иллюстрирует адаптивную ценность стратегии HDT в условиях высокой полузасушливости окружающей среды (Tuba et al. 1996). Лабораторные исследования показывают, что T. ruralis не изменяет содержание хлорофилла или каротиноидов во время циклов осушения и обезвоживания. (Tuba et al. 1996). Таким образом, снижение FV/Fm, наблюдаемое при изменении состояния воды по альбедо, указывает на обратимое физиологическое снижение общей способности к фотохимическому преобразованию света в высушенном состоянии и немедленное достижение устойчивого квантового выхода до достижения положительного прироста углерода во время регидратации, несмотря на широко изменяющиеся условия освещения. Координация Fv/Fm с закрытием побега, отраженная в измененном альбедо (Рис.4), предполагает, что эти растения обладают скоординированным набором анатомических и физиологических механизмов, вероятно, направленных на поддержание интегрированного ежедневного углекислого газа, необходимого для поддержания популяций в сильно изменяющихся засушливых и полузасушливых средах (Alpert & Oechel 1985).
Связь между Fv/Fm и альбедо была измерена в широком диапазоне суточных условий света, температуры и влажности окружающей среды, которые, вероятно, влияют на восстановление фотосинтеза (Deltoro et al. 1998b; Csintalan et al. 1999). В результате связь между альбедо и qP, NPQ или φPSII оказалась не такой тесной, как связь между этими параметрами и уровнем освещенности (рис. 5). В отличие от результатов исследований, в которых растения HDT сушили в контролируемых условиях (Eickmeier et al. 1993; Deltoro et al. 1998b; Csintalan et al. 1999), способность устанавливать NPQ не всегда повышалась в условиях сушки, а иногда снижалась при суточной сушке (Рис.5). Однако полученные нами результаты не касаются реакции NPQ на сушку как таковую, а скорее иллюстрируют способность устанавливать NPQ при различном освещении при регидратации после различных наборов предшествующих условий. Исследования сосудистых растений показывают, что давление возбуждения от первичного акцептора электронов Qa (оцениваемое по 1qP) в значительной степени не зависит от NPQ (Osmond 1994). Это имело место у T. ruralis, где сходные значения qP сопровождались очень разными кривыми светоотдачи NPQ после регидратации (Рис.5; см. также Csintalan et al. 1999). При длительных и благоприятных условиях влажности и освещенности, таких как 17 июня, Fv/Fm был высоким (рис.2), NPQ часто был высоким на всех уровнях освещенности, а φPSII демонстрировал незначительные суточные изменения после регидратации (Рис. 5). При высоких радиационных нагрузках, усиливающих быстрое утреннее высыхание (Рис.1, 19 июня), Fv/Fm, NPQ и φpsii сильно снижались при регидратации (Рис. 2 и 5).
Есть данные, что T. ruralis обладает признаками теневыносливого растения. (Tuba et al. 1998). У адаптированных к тени сосудистых растений ограниченные пулы ксантофилла и сниженная способность к обороту белка D1 приводят к накоплению нефункциональных реакционных центров PSII. Это служит для усиления теплового рассеяния избыточной световой энергии от неповрежденных реакционных центров, но не так быстро обратимо, как фотозащитное снижение эффективности фотосинтеза, опосредованное рН и циклом ксантофилла, которые характерны для солнечных растений (Demming-Adams & Adams 1992; Osmond & Grace 1994; Anderson et al. 1997). Таким образом, полуденное увеличение NPQ 17 июня могло быть результатом устойчивого NPQ после относительно короткого периода высокого PPFD, непосредственно предшествующего этим измерениям. Однако более высокие значения NPQ не сопровождались заметным снижением Fv/Fm (Рис.2), что говорит о том, что реакционные центры PSII были полностью функциональны. Более вероятным объяснением этого может быть то, что длительное раскрытие побегов, как указывает альбедо, приводило к большему интегральному PPFD в течение дня по сравнению с днями, когда закрытие побегов и обезвоживание уменьшали бы интегральный PPFD во время физиологической активности (Рис.2). Длительное воздействие света, вероятно, стимулировало циклический рост ксантофилла, связанный с рН, тем самым увеличивая NPQ в этих условиях. С другой стороны, высокие уровни освещенности, которые увеличивали NPQ во время быстрой сушки 19 июня, возможно, привели к более длительному и хроническому снижению фотохимической эффективности (Anderson et al. 1997). Это подтверждается результатами от 14 июня, когда PPFD был заметно ниже в утренние периоды, чем 19 июня. 14 июня Fv/Fm показал способность увеличиваться с регидратацией в течение дня (рис.2). В этих условиях NPQ оставался достаточно постоянным в течение дня (Рисунок 5). Эти результаты согласуются с предыдущими исследованиями, предполагающими, что высокоустойчивые к высыханию бриофиты поддерживают быстро реверсивные регуляторные фотозащитные механизмы в высушенном состоянии, которые могут быть адаптивными в условиях непредсказуемого водоснабжения (Deltoro et al. 1998b; Csintalan et al. 1999).
Таким образом, мы обнаружили, что толерантный к высыханию мох T. ruralis демонстрирует изменения в структуре всего растения при суточных колебаниях водной среды. Эти изменения сопровождались изменением максимальной фотохимической эффективности (Рис.4). Эти находки показывают, что устойчивые к высыханию бриофиты могут извлечь выгоду из воздействия низких уровней освещенности либо тогда, когда растения уже полностью увлажнены и восстановили максимальную чистую фотосинтетическую способность(Alpert & Oechel 1985; Tuba et al. 1996), либо при регидратации и во время декодирования от предыдущего высыхания (Casper et al. 1993; Csintalan et al. 1999). Кроме того, наши результаты показывают, что механизмы рассеивания световой энергии координируются с архитектурной реакцией на изменение водообеспеченности в установках HDT.
Благоданость
Это исследование было поддержано грантом НАТО Linkage Grant (ENVIR.LG 960897) г. Хенебри и Д. Гудину, а также американско-венгерской программой TeT (W/28/99) и OTKA 032568, предоставленной Z. Tuba.
Таблица 1. Значения F-критерия из двухстороннего ANOVA (Fv/Fm) и повторных измерений двухстороннего ANOVA (параметры гашения) параметров внутренней и внутрисуточной флуоресценции Tortula ruralis. Значения, отмеченные звездочкой, соответствуют p ≤ 0,05.
| Эффект | Fv/Fm | qP | NPQ | φPSII |
| Дата | 8.96∗ | 1.29 | 3.09∗ | 3.00∗ |
| Время | 9.53∗ | 5.79∗ | 0.34 | 7.35∗ |
| Дата × Время | 1.88 | 2.08∗ | 1.76∗ | 2.07∗ |
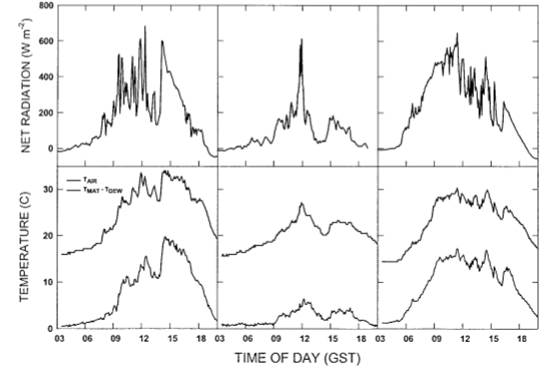
Рисунок 1. Суточное изменение (верхней) чистой радиации, полученной матрицей Tortula ruralis, и (нижней) температуры воздуха и отклонения от температуры точки росы мохового мата в течение трех дней исследования.
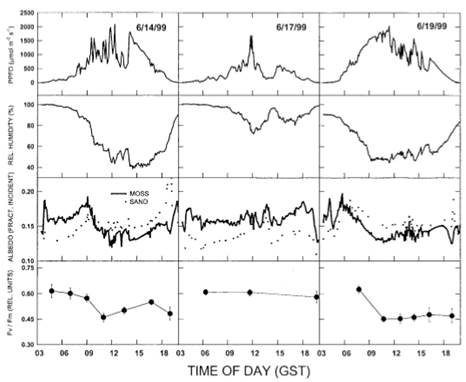
Рисунок 2. Суточное изменение (вверху) плотности фотосинтетического потока фотонов (PPFD), (второй ряд) относительной влажности, (третий ряд) альбедо песка и мохового ковра и (внизу) максимального фотосинтетического квантового выхода (Fv/Fm) T. ruralis в течение трех дней исследования. Каждая точка Fv/Fm - это среднее значение пяти измерений, столбики указывают ± один S.E. от среднего значения.
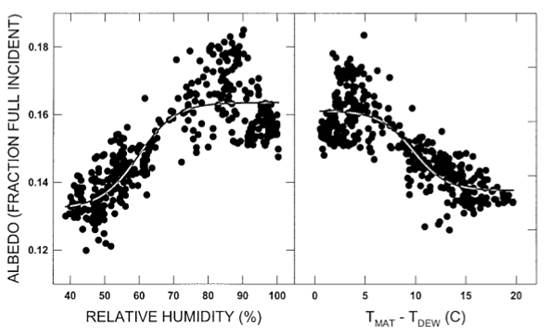
Рисунок 3. Соотношение между альбедо мохового ковра и (слева) относительной влажностью или (справа) отклонением мохового ковра от температуры точки росы.
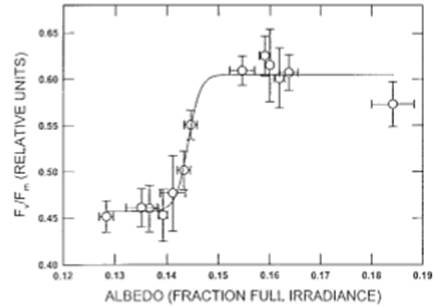
Рисунок 4. Отношение между альбедо и среднего Fv/Fm. Каждая точка представляет собой среднее из пяти измерений Fv/Fm и трех объединенных измерений альбедо (альбедо за пять минут до, в и через пять минут после времени темновой адаптации для Fv/Fm). Столбцы указывают ± 1 S.E. от среднего.
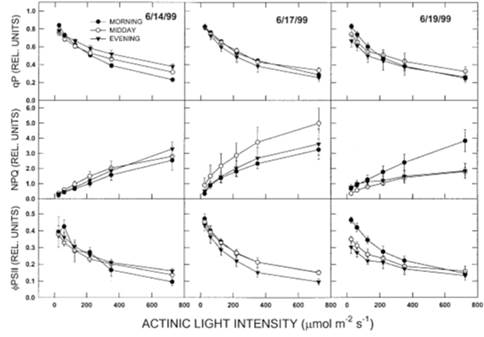
Рисунок 5. Суточное восстановление световых реакций фотохимического тушения флуоресценции qP (верхний ряд); нерадиационного тепловыделения NPQ (средний ряд); и эффективного фотохимического выхода PSII, φPSII (нижний ряд) Tortula ruralis в течение трех дней исследования. Каждая точка является средним значением пяти измерений; столбики указывают на ±1 S.E. от среднего значения.








