2020-08-05
2020-08-05 182
182Министерство образования и науки Российской Федерации
Сибирский федеральный университет
М.И. Гладышев
ЭКОЛОГИЧЕСКАЯ БИОФИЗИКА ВОДНЫХ СИСТЕМ:
КОНСПЕКТ ЛЕКЦИЙ
Красноярск
2013
СФУ
УДК 574.5(07)
ББК 28.072,1я73
Гладышев М.И. Экологическая биофизика водных экосистем: конспект лекций – Красноярск: Сибирский федеральный университет, 2013 – 79 с.
Настоящее издание является частью учебно-методического комплекса по дисциплине «Экологическая биофизика водных систем», включающее рабочую программу, конспект лекций, практикум, учебно-методические указания для самостоятельной работы.
Конспект лекций по курсу «Экологическая биофизика водных систем» рассматривает основные понятия, биофизические методы, экологические проблемы водоемов и способы их решения. В конце предлагается библиографический список.
Учебно-методический комплекс дисциплины «Экологическая биофизика водных систем» предназначен для магистров направления 020400.68 «Биология» магистерской программы «Гидробиология и ихтиология» - 020400.68.04.
УДК 574.5(07)
ББК 28.072,1я73
© Сибирский
федеральный
университет, 2013
СОДЕРЖАНИЕ
| Введение | 4 |
| Лекция 1. Введение: предмет и задачи экологической биофизики. Общие принципы и понятия экологии, используемые в экологической биофизике | 7 |
| Лекция 2. Биофизические методы мониторинга: Зондирование полей биолюминисценции в морях и полей флюоресценции в пресных водах. Интерпретация кривых вертикального распределения сигналов флюоресценции и биолюминесценции в терминах интегральных функциональных характеристик экосистемы | 21 |
| Лекция 3. Понятие о статистическом спектральном анализе. Математическое моделирование: Статистические и динамические модели. Простейшая динамическая модель на примере хемостата. Основные понятия: вектор состояния, параметры, константы. Уравнение роста в динамической модели | 38 |
| Лекция 4. Формальная теория сосуществования и регуляции видового состава сообщества. Принцип аутостабилизации и теорема об экологическом квантовании, их значение для поиска лимитирующих рост факторов в природных водоемах. Формализация понятия "качество воды" в динамических моделях. Выбор вектора состояния. Определение констант. Измерение фоновых концентраций для верификации модели | 53 |
| Лекция 5. "Распадные" модели динамики загрязняющих веществ, коэффициенты самоочищения. Некоторые проблемы гидробиологии, связанные с эколого-математическим моделированием качества воды | 62 |
| Лекция 6. Физическое моделирование: Типы МЭС и цели экспериментов с ними. Особенности экспериментального применения МЭС различных типов. Проблема адекватности МЭС и нативной экосистемы. "Эффект стенок" | 67 |
| Лекция 7. Особенности определения коэффициентов самоочищения легкоокисляемых поллютантов в МЭС | 75 |
| Лекция 8. Роль гидробионтов в физических процессах экосистемного масштаба на примере поверхностной пленки воды: Теплообмен и газообмен между водоемом и атмосферой. Пленки природных и антропогенных поверхностно-активных веществ (ПАВ) на воде. Влияние гидробионтов на тепло- и газообмен между водоемом и атмосферой. Информационное значение состава пленок природных ПАВ для мониторинга | 77 |
| Библиографический список | 97 |
ВВЕДЕНИЕ
В курсе «Экологическая биофизика водных систем» изучаются теоретические и методические основы экологической биофизики, рассматривающей физические процессы и явления, возникающие в результате функционирования живых надорганизменных систем, то есть даются основы мультидисциплинарного взаимодействия в области экологии. Данный курс знакомит студентов с теорией и практикой непосредственного применения комплексных (биологических, физико-математических и др.) методов при исследовании природных экосистем. Дисциплина предназначена для подготовки магистров по авторской программе «Гидробиология и ихтиология» по направлению 020400.68 – Биология (магистр) в соответствии с Федеральным государственным образовательным стандартом высшего профессионального образования.
Целью преподавания данной дисциплины является формирование у студентов базовых представлений об экологии как науке, об окружающей среде на основе использования новых физико-математических методов в экологии.
В результате изучения данной дисциплины обучающийся должен приобрести следующие практические навыки, умения, общекультурные (ОК), профессиональные (ПК) и специальные (СК) компетенции:
ОК-2: способен к инновационной деятельности;
ОК-4: понимает пути развития и перспективы сохранения цивилизации, связь геополитических и биосферных процессов, проявляет активную жизненную позицию, используя профессиональные знания;
ОК-6: способен самостоятельно приобретать с помощью информационных технологий и использовать в практической деятельности новые знания и умения, в том числе в новых областях знаний, непосредственно не связанных со сферой деятельности.
ПК-1: понимает современные проблемы биологии и использует фундаментальные биологические представления в сфере профессиональной деятельности для постановки и решения новых задач.
ПК-2: знает и использует основные теории, концепции и принципы в избранной области деятельности, способен к системному мышлению.
ПК-3: самостоятельно анализирует имеющуюся информацию, выявляет фундаментальные проблемы, ставит задачу и выполняет полевые, лабораторные биологические исследования при решении конкретных задач по специализации с использованием современной аппаратуры и вычислительных средств, демонстрирует ответственность за качество работ и научную достоверность результатов.
ПК-6: творчески применяет современные компьютерные технологии при сборе, хранении, обработке, анализе и передаче биологической информации.
ПК-9: профессионально оформляет, представляет и докладывает результаты научно-исследовательских и производственно-технологических работ по утвержденным формам.
В результате изучения дисциплины студент должен:
знать: структуру и функции природных экосистем как основу рациональной эксплуатации и охраны окружающей среды; роль живых организмов в гидрофизических процессах экосистемного масштаба;
уметь использовать методы мониторинга экосистем на основании зондирования биофизических полей биолюминесценции и флуоресценции;
владеть: методами математического и физического моделирования.
В связи с тем, что лекционный курс согласноФедеральному государственному образовательному стандарту высшего профессионального образования не должен превышать трети от всех аудиторных часов, то часть лекционного материала, помеченная * предназначена для самостоятельного изучения. Таким образом, сохранена структура курса и логичность изложения материала.
Лекция 1. Введение: предмет и задачи экологической биофизики. Общие принципы и понятия экологии, используемые в экологической биофизике*
В.И.Вернадский (1978) писал в 20-е годы, что человек как масштаб явлений бесконечно мал по сравнению с живой природой и легко подходит к свойствам ее элементов - организмов, но лишь путем трудной и долгой абстракции мы можем подняться до понимания свойств их совокупности - живого вещества. Таким образом, осознание необходимости системного подхода в экологии произошло уже давно, фактически с момента зарождения ее как самостоятельной науки, отличающейся от ботаники и зоологии. Но если теоретико-философские представления о целостности экологических систем, их интегральном функционировании, достаточно развиты, то в области практических исследований дела обстоят менее успешно. Как измерить интегральные функции экосистем, изучить их в воспроизводимом эксперименте и формализовать прогностических и оптимизационных математических моделях?
Для достижения подобных целей необходима специальная аппаратура, работающая с объектами, масштабы которых существенно больше масштабов индивидуального человеческого восприятия и методы практической абстракции: физическое и математическое моделирование. Именно это и является областью биофизики экосистем, как научной дисциплины, изучающей физические процессы, происходящие в живых надорганизменных системах и физические явления, вызванные функционированием данных систем, и разрабатывающей на этой основе методы изучения надорганизменных систем. Как же экологическая биофизика соотносится с экологией вообще и с гидроэкологией (гидробиологией) в частности? Поскольку объектами исследований экологической биофизики являются живые надорганизменные системы, то очевидно, что экологическая биофизика является составной частью экологии (гидробиологии), дополняющей классические биологические методы исследований (наблюдение, сравнительный анализ и др.) физическими подходами, заключающимися в аппаратурных измерениях изучаемых явлений, активных экспериментах в контролируемых условиях и математическом описании изучаемых объектов и явлений.
Таким образом, задачи и предмет экологической биофизики состоят в следующем.
1. Изучение физических процессов, являющихся отражением интегральных свойств экосистем и создание соответствующих методов мониторинга (по образному выражению академика И.И.Гительзона - "макроскопов").
2. Осуществление практической абстракции - физическое (создание экспериментальных экосистем) и математическое моделирование.
3. Изучение роли живых организмов в физических процессах экосистемного масштаба. (В отличие от традиционной биофизики, изучающей физические процессы в живых организмах, клетках, биомолекулах).
Представления об экологической биофизике были заложены в начале 70-х годов академиками И.А. Тесковым и И.И. Гительзоном, работавшими в отделе биофизики Института физики им. Л.В. Киренского Сибирского Отделения АН СССР (г. Красноярск). Логическим развитием работ по управлению популяциями микроорганизмов были исследования сообществ как одноклеточных, так и многоклеточных организмов в управляемых экспериментальных условиях. Венцом данного направления явилось создание в конце семидесятые годы замкнутой экологической системы жизнеобеспечения человека (ЗЭС - СЖО). Эта система была предназначена для обеспечения жизнедеятельности человека в условиях длительного космического полета и была поддержана руководителем космической программы СССР - академиком С.П. Королевым. В системе последнего поколения "Биос-3" (герметичном объеме около 300 м3) замкнутость по массе достигла 90 % (практически полное замыкание по газу и воде, вне круговорота - мясной рацион и твердые отходы). Экипажи из двух или трех испытателей жили в системе в течение 4-6 месяцев. За счет функционирования специально подобранных биотических звеньев (продуценты - консументы - редуценты) обеспечивался не только круговорот веществ, но и поддерживался постоянный газовый состав атмосферы. Таким образом, была создана первая (и до сих пор - единственная) в мире действующая модель биосферы Земли, включающая человека. Необходимо подчеркнуть, что популяции и сообщества, обеспечивающие круговорот, подбирались специальным образом, на основании точного расчета и знания их кинетических характеристик, полученных из ранее проведенных экспериментов по управляемому биосинтезу. Спустя 20 лет, в начале 90-х ученые США попытались создать аналогичную систему - "Биосферу-2", но эксперимент закончился неудачей: самоорганизации круговорота не произошло, резко изменился газовый состав воздуха и экипаж совершил аварийный выход из системы. Сопоставление двух систем является наглядным практическим подтверждением правильности эколого-биофизического подхода, согласно которому все эксперименты идут в строго контролируемых условиях, известен видовой состав основных звеньев, количественно измерены их кинетические характеристики, рассчитан коэффициент замыкания для данного объема и т.п.
Следует также отметить, что термин "экологическая биофизика" был введен И.А. Терсковым и И.И. Гительзон за два десятка лет до моды, возникшей на слово "экология", которое сейчас широко, порой - излишне, используется и в прессе, и в официальных документах. Наряду с академиками И.А. Терсковым и И.И. Гительзоном большой вклад в становление экологической биофизики внесли и другие выдающиеся ученые. Среди них один из главных создателей теории и практики создания замкнутых экологических систем проф. Б.Г. Ковров, один из основателей биоспектрофотометрии проф. Ф.Я. Сидько, основатель оригинального раздела математической экологии проф. Н.С. Абросов и другие.
Завершая краткое рассмотрение предметной и исторической связи экологической биофизики с другими разделами биологии, интересно провести некоторую аналогию с первым, состоявшимся более двухсот лет назад переходом естествоиспытателей в иной, несоизмеримый с человеком и его непосредственным восприятием масштаб изучаемых объектов. Это было проникновение в мир микроорганизмов. Оно состоялось благодаря изобретению специальной аппаратуры - микроскопов и проведению особых химико-биологических экспериментов. Следует отметить, что первоначально новая наука развивалась без участия представителей классической биологии. У ее истоков стояли шлифовальщик стекол (по современным понятиям - физик-оптик) А. Левенгук, химик Л. Пастер, врач Р. Кох. Более ста лет биологи отказывались от изучения новых объектов, К. Линней не включил микробов в свою систему классификации видов и отнес их к миру Хаоса. И лишь после длительного латентного периода возникла наука, именуемая микробиологией, без которой просто немыслима современная система представления о живой форме материи.
Таким образом, нет ничего удивительного, что многие процессы и явления, присущие надорганизменным системам, первыми увидели и исследовали не биологи (экологи), а именно физики, создавшие биофизические "макроскопы". Тем не менее, методы и подходы экологической биофизики, в частности - в области исследования водных систем, уже воспринимаются биологами (гидробиологами), и наряду с поколениями биофизиков-физиков новое поколение биофизиков-биологов, будет хорошо ориентироваться как в классической биологии, так и в гидрофизике, физическом и математическом моделировании.
В современной экологии существуют два основных подхода: "популяционный" и "экосистемный". В рамках первого подхода внимание исследователей сосредоточено на изучении особей того или иного вида, их взаимоотношений между собой, окружающей средой и другими видами. В рамках "экосистемного" или "функционального" направления изучаются интегральные процессы превращения вещества и энергии в результате взаимодействий абиотических и биотических компонентов надорганизменных систем. "Экосистемно-функциональный" подход в своей крайней форме утверждает, что трансформация энергии и циркуляция веществ, осуществляемые биологическими объектами важнее для исследования, чем сами объекты. Наряду с различием в подходах, существуют и разночтения в понимании терминов "популяция", "экосистема", "биогеоценоз" и ряда других.
Мы предварительно обсудим данные термины, используемые при изложении основ экологической биофизики. Прежде всего отметим, что "экосистема" и "биогеоценоз" будут использоваться как синонимы. Близкое значение этих понятий отмечали и крупнейшие зарубежные экологи (Одум, 1975), и сам автор термина "биогеоценоз" В.Н.Сукачев (1960). Основное различие состоит в том, что биогеоценоз имеет относительно четкие геоморфологические или гидрофизические границы, тогда как экосистемой могут называться системы любого ранга - от небольшого участка прибрежной полосы до Мирового Океана. Поэтому употребление термина "экосистема", как менее строгого и более универсального часто оказывается удобным и оправданным при общетеоретических рассуждениях. Следует также отметить, что более ранний и менее определенный термин В.И. Вернадского "живое вещество" по сути также весьма близок к понятию "экосистема".
Экосистемы (биогеоценозы) являются надорганизменными системами, поэтому их теоретический анализ логично проводить в терминах общей теории систем. Почему возникает необходимость в системном подходе и чем системы отличаются от "несистем"? Классический научный подход (называемый также аналитическим, физикалистским) характеризуется тем, что изучаются свойства отдельных элементов объекта, а затем из комбинации этих свойств дедуктивно выводятся свойства целого объекта. Совершенно очевидно, что для познания и практического использования многих объектов и явлений материального мира такой подход был и остается вполне адекватным и достаточным. Вместе с тем в ходе развития науки постепенно выявились объекты, которые не познавались традиционным способом. Например, по образному замечанию выдающегося нейрофизиолога, автора теории функциональных систем академика П.К. Анохина (1978), число возможных взаимодействий между клетками мозга выражается даже не астрономическим, а гораздо большим числом. То же самое можно сказать и о связях между организмами и абиотическими факторами в экосистеме.
Необходимость в системном подходе возникает тогда, когда невозможно получить адекватные знания об объекте на основании рассмотрения всех комбинаций взаимодействия между его элементами и когда невозможно создать модель объекта, которая была бы существенно меньше его самого. Следовательно, системы, также как и "несистемы", являются реальными объектами материального мира, и с этой точки зрения между ними нет материальных различий. Специфика систем лежит не в их природе, а в их отношении к познающему субъекту, то есть в форме получаемого о них знания и в способе представления этого знания. Такие знания базируются на представлениях об основных свойствах систем: целостности, упорядоченности, иерархичности. Рассмотрим эти категории на примере надорганизменных систем.
Основополагающее значение имеют работы автора учения о биосфере В.И. Вернадского, автора термина "экосистема" Э. Тенсли и автора учения о биогеоценозе В.Н. Сукачева. Не останавливаясь на различиях концепций (в том числе и терминологических) рассмотрим выявленные ими общие свойства данной системы.
Целостность означает несводимость свойств системы к сумме свойств составляющих ее элементов. Это положение четко прослеживается в концепциях надорганизменных систем: "Свойства живого вещества отнюдь не являются теми свойствами, которые мы изучаем при исследовании отдельного организма. В совокупности организмов - живом веществе - проявляются новые свойства, не заметные или не существующие, если мы станем изучать отдельный организм" (Вернадский, 1978, с. 52). "Биогеоценоз со своими закономерностями развития не есть просто сумма его компонентов и закономерностей их развития. Биогеоценоз в целом имеет свою качественную характеристику" (Сукачев, 1960, с. 9). Во-первых, из категории целостности следует, что не система определяется на основе определения элементов, а определение элементов выводится из определения системы. То есть при исследовании конкретного объекта как системы прежде всего должна быть определена концепция самой системы, и лишь затем, на основе этой концепции и вследствие ее возможно деление данной системы на элементы. Во-вторых, элементами системы могут быть объекты любой природы при том единственном условии, что для данной системы элементы остаются неделимыми единицами, причем их неделимость относительна и существует только в рамках данной системы. В третьих, для выдвижения концепции целостной системы необходимо определить ту объективную основу, те системообразующие связи,благодаря которым элементы (объекты различной природы) интегрируются в единое целое.
По определению В.Н. Сукачева "... биогеоценозом мы называем всякий конкретный участок земной поверхности, на протяжении которого сохраняется определенная система взаимодействия всех компонентов живой (растительность, животный мир и мир микроорганизмов) и мертвой природы (литосферы, атмосферы и гидросферы), т. е., иными словами, сохраняется однородной система превращения вещества и энергии и обмена ими с соседними биогеоценозами и другими телами природы" (с. 5). Очевидно, что элементами такой системы являются тела и вещества как живой, так и неживой природы. Это центральное положение концепции Э. Тенсли (Tansley, 1935): "В экосистеме организмы и неорганические факторы одинаково являются компонентами, которые находятся в относительно стабильном динамическом равновесии" (с. 306). Понятно, что системообразующей связью между элементами данной системы является обмен веществом и энергией.
Упорядоченность элементов экосистемы как результат ее функционирования. "Большей частью термин "система" употребляется там, где речь идет о чем-то собранном вместе, упорядоченном, организованном, но, как правило, не упоминается критерий, по которому компоненты собраны, упорядочены организованы" (Анохин, 1978, с. 53). Согласно представлениям П.К.Анохина, фактором, упорядочивающим структуру системы, ее функционирование, является результат деятельности системы, который оказывает на систему влияние по принципу обратной связи, избавляет ее элементы от избыточных степеней свободы, детерминирует взаимодействие элементов системы. Не может быть понятия системы без ее полезного результата. Действительно, понимание системы как множества любых элементов, между которыми возможны какие угодно взаимодействия, не приводящие к некоторому результату, описываемому достаточно компактно, гносеологически бессмысленно, так как такая "система" в принципе не может быть исследована и познана. Мы не имеем полного знания о всех особенностях структуры и функционирования системы, взаимосвязях всех элементов, но результат должен быть вполне представим и носить конкретный характер. Таким образом, вся деятельность системы и ее изменения должны быть представлены в терминах результата.
По мнению В.Н. Сукачева, почва как компонент биогеоценоза является и суммарным результатом взаимодействия всех его компонентов, т.е. почва в известном смысле - итог процессов, происходящих в биогеоценозе. Для водных экосистем аналогичным результатом можно назвать комплекс растворенных и взвешенных в воде веществ (химическое качество природных вод) и донные отложения. Кроме того, результатом функционирования как наземных, так и водных экосистем, кроме почвы и качества воды, является присущий им комплекс организмов - живое вещество или видовой состав экосистемы (понятие "видовой состав экосистемы" будет обсуждаться ниже).
Очевидно, что состав среды (почвы, воды) и организмов, складывающийся в результате функционирования экосистемы, влияет на само функционирование, изменяет его и вследствие этого сам подвергается соответствующим изменениям. Поэтому представления В.Н. Сукачева о результате функционирования биогеоценоза согласуются с концепцией П.К. Анохина об упорядочивающем результате функционирования системы. Конкретному типу почвы всегда соответствует растительность, но и тип почвы складывается в результате взаимодействия организмов и среды. Аналогию можно провести и для водных систем.
Иерархичность экосистемы. Иерархичность системы выражается в том, что каждый ее элемент является системой, а она сама - элементом системы высшего уровня. Однако это не означает, что материальный мир представляет собой линейную последовательность подсистем и надсистем, вложенных одна в другую. Один и тот же объект, обладающий многоплановой качественной спецификой, может иметь с окружающим материальным миром разносторонние системообразующие связи, т.е. одновременно быть элементом разных систем, не соподчиненных иерархически. "Мы можем говорить о некотором множестве элементов как о системе лишь относительно определенных свойств и отношений этих элементов " (Садовский, 1974, с. 185).
Согласно концепциям В. И. Вернадского и В. Н. Сукачева, биогеоценозы как подсистемы являются элементами биосферы, представляющей по отношению к ним иерархически высшую надсистему. Аналогично понимал экосистемы Э. Тенсли, называя их основными единицами природы на поверхности Земли. В настоящее время эти взгляды разделяют практически все экологи.
В то же время вопрос об элементах экосистемы нельзя назвать решенным. Широкое распространение имеет следующая схема линейной иерархии биологических систем:... ® клетка ® организм ® популяция ® экосистема
То есть считается, что популяции - это элементы экосистем. В связи с этим кратко рассмотрим популяцию как биологическую систему.
Популяция как надорганизменная система В настоящее время существует два подхода к определению популяции: 1) "генетический" и 2) "экологический. Определим их основные особенности.
1. "Популяция - это минимальная самовоспроизводящаяся группа особей одного вида, на протяжении эволюционно длительного времени населяющая определенное пространство, образующая самостоятельную генетическую систему и формирующая собственное экологическое гиперпространство" (Яблоков, 1987, с. 150). Под системообразующей связью, интегрирующей совокупность особей в единое целое подразумевается обмен генами в процессе полового размножения, а под элементом системы - особь, как носитель определенного генотипа.
2. "Популяция есть население, образующее относительно простые или иерархически сложные скопления, функционирующие как звено цепи питания, регулирующие свою продуктивность в соответствии с продуктивностью других звеньев этой цепи и продуцирующие видоспецифическое живое вещество" (Межжерин, 1975). Здесь понятие популяции основано на ее трофике, т.е. полагается, что целостной системой популяцию делают в первую очередь схожие трофические функции ее особей.
Группа особей одного вида действительно имеет сходную трофическую функцию, сходную морфологию особей и т.п. Однако, общность питания и строения - это не реальные связи, интегрирующие особей в единое целое, а лишь признаки принадлежности их к одному виду. Спектр питания популяции - это именно сумма конкретных трофических функций (отнюдь не идентичных) ее особей. "Нет ни одного вида животных, у которых возрастные группы не обладали бы разными спектрами питания, Очень часто существенно различаются в питании половые группы" (Яблоков, 1988, с. 92). В то же время в природе широко распространена конкуренция за ресурсы питания, т.е. спектры питания разных видов организмов перекрываются. И если системообразующая связь между особями популяции - это питание одним видом пищи, то, во-первых, точно такая же связь существует в экосистеме между особями разных видов, во-вторых, подобная связь отсутствует между особями одного вида, но разного возраста, стадии или пола. Таким образом, наиболее обоснованным с биологической точки зрения и с позиций общей теории систем выглядит положение, что системообразующей связью популяции является обмен генами между ее особями, происходящий в процессе полового размножения. Функционирование популяции как целостной системы, обладающей только ей присущими качествами, проявляется в эволюционном процессе.
Определение популяции как реальной системы, основанной на генетических связях, отнюдь не отрицает возможности выделения ее в конкретных исследованиях на основе экологических и морфологических признаков ее особей. При выделении популяции ведущими критериями могут оказаться именно экологические и фенетические, а не генетические признаки. Это не противоречит одно другому: пренебрежение как теоретическими определениями, так и практическими критериями выделения объекта, абсолютизация одного из них одинаково вредны для процесса познания.
Понятно, что элементом популяции является особь. Основным свойством особей как элементов популяции является наличие у них определенных генотипов, различных по составу генов. Очевидно, что это свойство особи не зависит от ее возрастной стадии и трофической функции.
Соподчиненность экосистемы и популяции. Как отмечалось выше, экосистема - это система интегрированная на основе обмена веществом и энергией между ее элементами. С данной точки зрения популяция не является неделимым элементом экосистемы. Подрост (торчки) древесных растений поедаются копытными животными, взрослые деревья - насекомыми. Мальки многих рыб являются планктофагами, тогда как взрослые особи - бентофаги или хищники. Во всех примерах особи составляют единую популяцию, но функционируют в качестве разных элементов экосистемы (биогеоценоза). Особь является как элементом популяции, так и элементом экосистемы, но благодаря разным, объективно присущим ей сторонам качества, поэтому понятия особи в популяции и экосистеме отражают разные системообразующие свойства данных объектов.
Если в популяции основным свойством особи является качество ее генотипа, вклад в эволюционный процесс, то в экосистеме - вклад в процессы превращения вещества и энергии. Как для минералов, так и для организмов имеют значение масса, состав и энергия живого вещества. Таким образом, реальным элементом экосистемы, наряду с телами и веществами неживой природы, является не популяция, а особь, характеризуемая как живое вещество. Видовой состав экосистемы - это состав живого вещества. Таким образом, представляется неверной постановка вопроса об иерархической подчиненности популяции экосистеме. Популяция и экосистема - это системы не разного уровня, а разного типа интеграции и не могут быть объединены в линейную систему иерархии. В. Н. Сукачев (1964) отмечал, что ставить биогеоценоз в общий ряд биологических явлений (... клеточный ® организменный ® популяционный ®...) не следует, "...биогеоценотический уровень изучения явлений природы не относится к серии биологических уровней, это уровень особого порядка".
Структура природы оказывается в конечном итоге предельно простой: она определяется двумя системами интеграции - видовой и биогеоценотической. Схему уровней организации природы, предлагаемую С.С.Шварцем; можно представить следующим образом:
Видовая (биологическая) система интеграции..Клетка ® организм ® популяция ® вид
¯
Биогеоценотическая система интеграции биогеоценоз
Мы разделяем взгляды С.С.Шварца, за исключением одного пункта: реальным элементом биогеоценоза является не популяция, а особь, благодаря наличию у нее, кроме специфических биологических (генетико-эволюционных) свойств, также и свойств геохимических, свойств живого вещества. Для иллюстрации концепции, используемой в данном курсе экологической биофизики, предлагается следующая схема:
Биологический тип интеграции ...Клетка ® особь ® популяция ® вид
(генотип)
¯
Биогеохимический тип интеграции Особь ®? ® экосистема ® биосфера
(живое вещество)
Необходимо подчеркнуть, что понятие особи как элемента экосистемы до конца не ясно как в теоретическом, так и в практическом плане. Например, агрегаты бактериальных клеток или микроводорослей в водоеме, муравейник, совместно охотящаяся пара млекопитающих, приносящая добычу детенышам и т.п., т.е. группы организмов, выполняющих "геохимическую работу" как единое целое, видимо, могут являться неделимыми единицами экосистемы. Что касается популяции, то в качестве неделимого элемента, обладающего только ему присущей целостной функцией (специфической эволюционной судьбой), она выступает в отношении вида. В экосистеме неделимость популяции и целостность ее функции "исчезают". Однако это "исчезновение" популяции в рамках биогеохимической системы интеграции следует понимать лишь в том смысле, что она не является элементом экосистемы. Экосистема выступает по отношению к популяции не как надсистема, а как среда.
Итак, экосистемы и популяции представляют собой надорганизменные системы разного типа, несводимые одна к другой. Их изучение имеет объективно различные конечные цели, выражаемые в терминах результата функционирования этих систем. Итогом исследования экосистемы должен быть прогноз (с целью управления и оптимизации) химического состава биотопа (почвы или воды) и состава живого вещества в зависимости от внешних воздействий: погодных условий, внешних потоков вещества и энергии (в том числе - антропогенных загрязнений). Что касается состава живого вещества, то есть видового состава экосистемы, то "вид" в экологической биофизике - это группа особей, имеющих одинаковые кинетические характеристики взаимодействия с веществом экосистемы. Как правило, внутри каждой популяции существует несколько таких групп. Кроме того, биологическое время функционирования популяций обычно длиннее астрономического времени функционирования экосистемы, т.е. за один вегетационный сезон (основная временная единица экосистемы) популяция не успевает осуществить несколько циклов обмена генами, поэтому микроэволюционный процесс на этом масштабе времени не срабатывает и, соответственно, не учитывается при изучении и моделировании функционирования экосистемы. Поэтому при дальнейшем изложении основ экологической биофизики мы будем иметь дело не с генетическими эволюционирующими популяциями, а с биогеохимическими "видами" - живым веществом.
Лекция 2. Биофизические методы мониторинга: Зондирование полей биолюминисценции в морях и полей флюоресценции в пресных водах. Интерпретация кривых вертикального распределения сигналов флюоресценции и биолюминесценции в терминах интегральных функциональных характеристик экосистемы
Жизнедеятельность сообществ и отдельных организмов вносит возмущения в физические поля: электрическое, световое и др. Сигналы, генерируемые биологическими объектами, могут быть зарегистрированы соответствующей физической аппаратурой и служить основой для изучения функционирования надорганизменных систем. Наиболее широко используемые для этой цели явления - биолюминесценция и флюоресценция. Прежде чем перейти к рассмотрению особенностей каждого из явлений, отметим их общие свойства.
Оптическое поле, создаваемое живыми излучателями, не может быть описано только физическими законами, характерными для гидрооптического поля, например, геометрическими законами ослабления. Например, в океане можно наблюдать такое явление, когда биолюминесцентная вспышка одного животного возбуждает излучение другого, и свет распространяется на многие километры без затухания. Поэтому представляется возможным говорить о существовании биофизических полей: поля биолюминесценции и флюоресценции, для которых характерны свои эколого-биофизические законы.
Второй характерной особенностью биолюминесценции и флюоресценции как экологических явлений является то обстоятельство, что для их изучения, как правило, не может быть использована стандартная гидрофизическая (гидрооптическая) аппаратура. Для исследования биофизических полей требуется создание специальной аппаратуры с особыми параметрами пространственно-временного разрешения и чувствительности.
Природа биолюминесценции и характеристика светящихся организмов. Светящиеся организмы обитают только на суше и в море, в пресных водах и соленых озерах их нет (хотя нельзя исключать, что где-нибудь в глубинах неисследованных соленых озер они когда-нибудь будут найдены). Поэтому, с точки зрения экологической биофизики водных систем, биолюминесценция - чисто морское явление, обнаруживаемое при солености от 10 0/00 и выше (знак 0/00 "промилле" означает количество граммов солей в 1000 г раствора). Поэтому, например, в Балтийском море, соленость которого около 2 - 8 0/00 , биолюминесценция отсутствует (за исключением более соленых глубинных слоев).
Биолюминесценция - это разновидность хемилюминесцентной реакции, т.е. реакции испускания оптического излучения за счет расходования энергии химических связей. Характерной особенностью биолюминесценции, выделяющей ее в ряду прочих хемилюминесцентных реакций, является обязательное участие в них специфических ферментов - люцифераз. У большинства морских организмов биолюминесцентная реакция протекает в две стадии. На первой стадии за счет расхода энергетических эквивалентов клетки, таких как АТФ или НАД×Н образуется (восстанавливается) субстрат ферментативной реакции, LH2 . У бактерий субстратом является флавинмононуклеотид (ФМН) и альдегид, у других организмов - люциферин. На второй стадии происходит ферментативное окисление субстрата кислородом. В результате реакции образуется квант света, hn. Характер свечения зависит от вида люциферазы, который является специфичным у разных видов организмов.
Различают внутриклеточное и внеклеточное свечение. При внутриклеточном свечении молекулярные фермент-субстратные системы локализованы внутри клеток. У многоклеточных животных свечение обеспечивается специальными клетками - фотоцидами. Иногда свечение многоклеточных организмов происходит за счет светящихся симбиотических бактерий. Внеклеточное свечение встречается у некоторых ракообразных. Они выбрасывают из специальных желез струи люциферина и люциферазы в окружающую среду, где при их смешении возникает свечение.
В настоящее время известно значительное количество видов светящихся бактерий. Среди них род Photobacterium, представители р. Vibrio и др. В отличие от всех остальных светящихся организмов бактерии излучают свет непрерывно. При высоких концентрациях бактерий в морской воде возникает так называемое "молочное свечение". Светящиеся бактерии могут расти и в анаэробных условиях, но светятся только при наличии в среде кислорода. Большинство светящихся бактерий являются симбионтами обитающими в кишечниках рыб, кальмаров и др., их пребывание в свободном состоянии в морской воде носит временный характер. Как считается, излучение света бактериями является своеобразной приманкой, обеспечивающей им возможность попасть из воды в кишечник хозяина. Хотя, как отмечалось, численность светящиеся бактерии в морской воде может быть высокой, в целом их вклад в формирование биолюминесцентного поля океана относительно невелик.
Основной вклад в биолюминисценцию пелагиали морей и океанов вносят одноклеточные панцирные жгутиконосцы - динофлагелляты, которые, как известно, одновременно относятся и к простейшим животным, и к водорослям (Dinophycea или Peridineae). Среди них наиболее известны ночесветки - виды р. Noctiluca, широко распространенные во многих морях, в том числе - в Черном. В открытом океане доминируют виды р. Pyrocystis, которые, в отличие от ночесветок, утративших хлорофилл, способны к фотосинтезу.
Второй по значимости группой организмов, формирующих биолюминесцентное поле океана, являются ракообразные: ракушковые (Ostracoda), веслоногие (Copepoda) и Euphauseacea. Особенно большой численности могут достигать остракоды, среди них - представители рода Cypridina. Люминесцентные вспышки ракообразных длятся от 0.15 до 3 с, латентный период между вспышками может продолжаться около 0.05 - 0.20 с. Как считается, люминесценция животных этой группы, как и других многоклеточных, может иметь сигнальное значение, служить для привлечения добычи и защиты от хищников.
Кроме перечисленных групп животных, биолюминесценция характерна для представителей радиолярий, кишечнополостных, кольчатых червей, моллюсков, некоторых иглокожих (за исключением морских ежей) и др. Также в настоящее время известно более 300 видов светящихся рыб, причем только около 50 видов светятся за счет симбиотических бактерий. Световая реакция живых организмов наступает в ответ на какое-либо раздражение: механическое, световое, ультразвуковое. Спектр биолюминесценции зависит от вида организма, однако у большинства из них основной максимум излучения находится в диапазоне длин волн 470 - 480 нм, то есть соответствует максимуму пропускания морской воды. Биомасса светящихся организмов может достигать 80% биомассы планктона, поэтому биолюминесценция является весьма информативной характеристикой морских сообществ, для изучения которой могут быть применены биофизические приборы.
Аппаратура для измерения морской биолюминесценции. Особенности аппаратуры и методов исследования биолюминесцентного поля связаны с тремя его основными свойствами: а). Импульсный характер излучения, который, в отличие от медленно изменяющегося астрономического света, требует высокого быстродействия аппаратуры; б) необходимость отделения световых сигналов живых организмов от астрономического фона; в) необходимость искусственного раздражения организмов для стимулирования биолюминесценции.
На основе перечисленных принципов были созданы разнообразные приборы для вертикального и горизонтального зондирования биолюминесцентного поля океана - погружаемые и буксируемые батифотометры. В России наиболее широко применялся комплекс вертикального зондирования "Ромашка". При погружении комплекса с борта судна на кабель-тросе осуществляется непрерывная запись сигнала биолюминесценции в воде, протекающей через рабочую камеру, где происходит механическое возбуждение организмов за счет повышенной турбулентности. Вертикальный профиль сигнала биолюминесценции в реальном времени отображается на мониторе, и по команде оператора, на горизонте, представляющем интерес, срабатывает один из шести батометров, входящих в комплекс, и берется реперная гидробиологическая проба. Весьма перспективной для мониторинга является также аппаратура типа "Факел". Его отличие от "Ромашки" заключается в том, что в рабочей камере расположены сразу три фотоприемника, распределенные по ее длине.
Изучение пространственно-временного распределения планктона по биолюминесценции. Первоначально внедрение биофизических зондов (в том числе - фотометров) в практику гидробиологических исследований было вызвано необходимостью получения точной картины пространственно-временного распределения биомассы гидробионтов. Как известно, гидробиологические методы сбора планктона весьма трудоемки и имеют чрезвычайно малую разрешающую способность в пространстве и времени. Это неизбежно приводит к ошибочным представлениям об однородном распределении и плавном изменении биомассы планктона в океанах.
Переход к отбору "точечных" батометрических проб несколько уточнил информацию о распределении планктона, но не смог кардинально ее улучшить. Интервалы между батометрическими точками в пространстве и времени по-прежнему оставались чрезвычайно большими, а самих "точек" бралось чрезвычайно мало. Как известно, камеральная обработка проб планктона производится визуально, под микроскопом, занимает много времени и требует весьма высокой квалификации и длительной подготовки специалиста, который должен в совершенстве знать систематику водорослей или животных. Один исследователь может обработать примерно сотню рутинных камеральных проб в год. По такому количеству точек невозможно уловить ни микромасштабную (метры и минуты), ни макромасштабную (акватории морей и океанов, месяцы) динамику изменчивости планктона. Очевидно, что батифотометры, способные регистрировать характеристики биолюминесцентного поля в непрерывном режиме, со скоростью движения судна или опускаемого троса, способны снять многие ограничения в изучении пространственно-временной структуры пелагических экосистем.
На первом этапе применения биофизических зондов необходимо было найти количественную зависимость между интенсивностью биолюминесценции и биомассой планктонных организмов. Очевидно, что коэффициенты пересчета определяются видовым составом планктона, поэтому их следует определять для каждого района и сезона. Тем не менее, видовой состав - довольно крупномасштабная и инерционная характеристика, поэтому число районов, в которых необходимо определять значения коэффициентов, сравнительно невелико. В конечном итоге, необходимые зависимости были получены. В результате появилась возможность по сигналу биолюминесценции определять биомассу.
Зондирование биолюминисцентного поля позволяет выявить точную картину распределения планктона в океане и исследовать зависимость этого распределения от факторов среды. На основании полученных данных можно сделать вывод, что для распределения планктона в океане характерна пространственно-временная гетерогенность различного масштаба. Неоднородности в распределении планктона зависят от гидрофизических и гидрохимических условий.
Наиболее часто наблюдается связь биолюминесценции с градиентом температуры воды. Водные массы различной температуры, как известно, имеют разную плотность. Именно этим обусловлено возникновение в пелагиали особых физических структур, например, термоклина. Именно в зоне термоклина, где кроме резкого градиента температуры наблюдаются градиенты концентраций биогенных элементов и др., часто расположен и максимум биолюминесценции, т.е., максимум биомассы планктона. Перемещение водных масс с различной температурой (плотностью) характеризует процесс образования внутренних волн, когда происходит периодический подъем к поверхности и опускание более холодных глубинных слоев. Имеются участки океана, например, вблизи берегов Перу, где происходит постоянный подъем холодных глубинных слоев, называемый апвеллингом. Поскольку глубинные воды богаты биогенными элементами, в районах их выхода на поверхность наблюдается высокая продукция и биомасса фитопланктона и зоопланктона.
Наряду с гидрофизическими неоднородностями, вызванными перемещениями водных масс различной температуры и плотности, гетерогенные структуры в поверхностном слое океана могут формироваться под действием ветра. На поверхности морей (а также крупных озер) часто возникает явление, называемое циркуляцией (или вихрями) Ленгмюра. При этом вода движется по замкнутым траекториям, образуя упорядоченные ячейки. На поверхности возникают зоны дивергенции (расхождения) и конвергенции (схождения) потоков воды. Ширина пенных полос в океане обычно от одного до нескольких метров, расстояние между ними - от 2 до 300 м (в озерах ширина полос - несколько десятков сантиметров, расстояние между ними - от 2 до 25 м). В зонах конвергенции скапливаются не только органические вещества (пена), но и планктонные организмы, отчетливо прослеживающиеся в структуре биолюминесцентного поля.
Наряду с неоднородностями распределения планктона, формирующимися в результате воздействия гидрофизических факторов, существуют и чисто экологические (биологические) причины образования скоплений гидробионтов. В олиготрофных районах океана, где концентрации элементов питания в среднем чрезвычайно низки, жизнь может существовать только в виде скоплений. Образование скопления может происходить следующим образом. В области появления локального пятна повышенных концентраций биогенных элементов (например, за счет энтрузии из глубинных слоев) развивается фитопланктон. Высокие концентрации микроводорослей, в свою очередь, обуславливают скопление и быстрый рост зоопланктона. Зоопланктон, поедая микроводоросли, обеспечивает тем самым рециркуляцию биогенных элементов. Выделяемые метаболиты способствуют росту бактерий, также осуществляющих процесс быстрой минерализации биогенов. Рециркуляция биогенных элементов способствует дальнейшему росту фитопланктона. Если скорость круговорота минеральных элементов и роста организмов выше скорости размывания образовавшегося скопления, то планктонное "облако" может существовать длительное время и самоподдерживаться за счет экологического круговорота.
Таким образом, в продуктивных районах океана формирование пространственных неоднородностей в биолюминесцентном поле (распределении планктона) связано с гидрофизическими факторами, тогда как в олиготрофных районах неоднородности в основном обусловлены биологическими механизмами. В настоящее время биофизические зонды выполняют две взаимосвязанных функции: 1) визуализация пространственного распределения планктона для прицельного отбора гидробиологических проб и изучения неоднородностей; 2) уточненная оценка биомассы планктона и ее пространственно-временного распределения.
Первая функция зондов заключается в том, что если ранее океанологи на станциях отбирали пробы фактически наугад, на стандартных "круглых" горизонтах, то теперь, получив за считанные минуты профиль биолюминесценции, специалисты выбирают на его основе наиболее интересные точки для отбора батометрических проб. Что же касается изучения неоднородностей в распределении планктона, то ранее, до применения биофизических зондов, их просто не видели из-за низкого пространственно-временного разрешения гидробиологических методов. В настоящее время неоднородности стали доступны для наблюдения и изучения. Очевидно, что более высокое пространственно-временное разрешение биофизических методов имеет решающее значение и для выполнения их второй функции по уточнению биомассы планктона и в локальном, и в глобальном масштабах. Так, в относительно короткие сроки была создана карта интенсивности биолюминесценции Мирового океана, отражающая глобальное распределение биомассы планктона.
Несомненно, информация о распределении биомассы гидробионтов, весьма важна для изучения состояния экосистем и контроля качества природной среды. Но биомасса является статической характеристикой, а как известно на основании учета только статических характеристик нельзя получить адекватных знаний об экосистеме. Для понимания процессов функционирования экосистем и прогноза их состояния необходимо уметь определять кинетические характеристики,т. е., скорости изменения биомассы гидробионтов и концентрации веществ. К кинетическим характеристикам относятся скорости роста, смертности, фотосинтеза, деградации поллютантов и т.д. Важнейшими кинетическими характеристиками являются продукция (A) и деструкция (R) и их соотношение (A / R), определяющее трофический статус водоема: евтрофный (A / R > 1), мезотрофный (A / R»1) или олиготрофный (A / R < 1).
Измерения интегральной продукции и деструкции в пелагических экосистемах производится методом светлых и темных склянок в кислородной или радиоуглеродной модификации. В отношении пространственно-временного разрешения этот метод обладает теми же недостатками, что и батометрические пробы. Поэтому важно было выяснить возможности применения биофизических зондов для определения кинетических характеристик водных экосистем.
Как выяснилось в результате многолетних исследований академика И.И.Гительзона с сотрудниками, форма кривой вертикального распределения биолюминесценции зависит от продуктивности района. Можно выделить 5 основных типов вертикального распределения интенсивности биолюминесцентного поля.
1. Равномерное (нестратифицированное) вертикальное распределение интенсивности биолюминесцентного поля характерно для олиготрофных районов. Как правило, интенсивность биолюминесценции таких районов низка.
2. Распределение интенсивности биолюминесценции с максимумом на поверхности наблюдается в прибрежных продуктивных районах, где ветры и течения, неоднократно сменяющие направления в течение суток, обеспечивают быстрое перемешивание всех слоев. В то же время солнечная радиация, поверхностный сток биогенных элементов и относительно высокая мутность воды создают предпосылки для интенсивного развития фитопланктона именно на поверхности. Биолюминесцентные сигналы фитопланктона и формируют поверхностный максимум.
3. Распределение с одним подповерхностным максимумом наблюдается в высокопродуктивных (евтрофных) районах океана, в зонах апвеллингов, то есть подъемов глубинных вод, богатых биогенными элементами. Как правило, максимум расположен на глубине 5 - 30 м в зоне термоклина и резкого изменения концентрации биогенных элементов. Такое расположение максимума жизнедеятельности планктоннного сообщества, индикатором которого является биолюминесценция, объясняется тем, что ниже этой зоны интенсивность развития фитопланктона сдерживается недостатком света, а выше - недостатком биогенных элементов.
4. Двухмаксимумное вертикальное распределение интенсивности биолюминесценции характерно для мезотрофных районов. Верхний максимум расположен на горизонте 20 - 50 м, нижний - на глубине 60 - 100 м. Верхний максимум биолюминесценции совпадает с максимумом фотосинтетической продуктивности.
5. Многомаксимумное вертикальное распределение интенсивности биолюминесцентного поля типично для районов со сложной гидрологией, т.е. с несколькими термоклинами или взаимопроникновением водных масс различного состава. Например, третий максимум на глубине 300 м характерен для районов Атлантики, в которые проникают ответвления течения Гольфстрим со своим планктонным населением.
Найденные закономерности позволяют в течение нескольких минут, по форме кривой вертикального распределения поля биолюминесценции, определять трофический статус района. В данном случае уже неважно, насколько сигналы зондирующей аппаратуры коррелируют с биомассой, важна сама форма кривой, регистрируемая биофизическим зондом. Таким образом, профиль вертикального распределения интенсивности биолюминесцентного поля является специфической эколого-биофизической характеристикой, несущей информацию не только о распределении отдельных групп организмов, но и об интегральном функционировании пелагической экосистемы.
Интерпретация распределения сигнала флюоресценции. При гидробиологических исследованиях экосистем внутренних водоемов возникают те же проблемы пространственно-временного разрешения применяемых методов, что и в океанах. Биолюминесценция в слабосоленых и пресных водах отсутствует. Основным объектом для эколого-биофизического зондирования является поле флюоресценции хлорофилла. К настоящему времени наиболее широкое распространение получили контактные методы регистрации флюоресценции.
Флюоресценция хлорофилла - это разновидность фотолюминесценции, то есть свечения веществ, источником энергии возбуждения которого являются поглощенные кванты света. При поглощении молекулой кванта света ее электрон (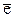 ) из основного (невозбужденного) синглетного состояния, S0, переходит на энергетический более высокий (возбужденный) синглетный уровень S*. Синглет-синглетный переход происходит без обращения спина электрона. Хлорофилл поглощает свет в синей и красной области. Как с синего, так и с красного возбужденного уровней электрон может отправиться в электрон-транспортную цепь (ЭТЦ) и участвовать в первичных фотохимических реакциях фотосинтеза. Второй путь для электрона - вернуться на основной уровень, отдав энергию в виде тепла и флюоресценции. При этом электрон с синего возбужденного уровня не может непосредственно вернуться на S0, он переходит сначала на красный уровень, рассеивая часть энергии в виде тепла. Флюоресценция происходит только при возвращении электрона с более низкого красного возбужденного уровня, поэтому флюоресценция хлорофилла проявляется только в виде красного света. Это обстоятельство является чрезвычайно важным для создания регистрирующей аппаратуры.
) из основного (невозбужденного) синглетного состояния, S0, переходит на энергетический более высокий (возбужденный) синглетный уровень S*. Синглет-синглетный переход происходит без обращения спина электрона. Хлорофилл поглощает свет в синей и красной области. Как с синего, так и с красного возбужденного уровней электрон может отправиться в электрон-транспортную цепь (ЭТЦ) и участвовать в первичных фотохимических реакциях фотосинтеза. Второй путь для электрона - вернуться на основной уровень, отдав энергию в виде тепла и флюоресценции. При этом электрон с синего возбужденного уровня не может непосредственно вернуться на S0, он переходит сначала на красный уровень, рассеивая часть энергии в виде тепла. Флюоресценция происходит только при возвращении электрона с более низкого красного возбужденного уровня, поэтому флюоресценция хлорофилла проявляется только в виде красного света. Это обстоятельство является чрезвычайно важным для создания регистрирующей аппаратуры.
Поскольку все синглет-синглетные переходы осуществляются без изменения спина электрона, флюоресценция происходит очень быстро: время жизни молекулы в возбужденном состоянии S* составляет порядка 10-8 - 10-9 с. Этим флюоресценция отличается от фосфоресценции - разновидности фотолюминесценции, при которой молекула излучает квант света при возвращении электрона с триплетного уровня, происходящем с изменением спина.
Для изучения флюоресценции фитопланктона создаются специальные приборы - флюориметры. Принцип действия флюориметров состоит в следующем. Кювета, содержащая пробу воды с исследуемыми микроводорослями, для возбуждения флюоресценции облучается через соответствующие светофильтры коротковолновым светом (сине-зеленая область). Перед регистрирующим устройством стоит красный светофильтр, который отрезает возбуждающий свет и пропускает только флюоресценцию. В зависимости от конструкции, флюориметры могут использоваться: а) в бортовом (лабораторном) варианте для точечных измерений; б) в бортовом варианте с проточной кюветой для непрерывной регистрации на ходу судна горизонтального распределения флюоресценции; в) в погружаемом варианте для регистрации вертикального распределения флюоресценции.
Необходимость применения флюоресцентных зондов в гидробиологии вызвана необходимостью получения точной картины пространственно-временного распределения фитопланктона. Поэтому на первом этапе решается задача, аналогичная задаче начального этапа биолюминесцентных исследований: определение количественной связи между интенсивностью флюоресценции (I фл) и концентрацией хлорофилла а фитопланктона (С хл). На удельный выход флюоресценции, R = I фл / С хл , влияет множество факторов: видовой состав фитопланктона, условия среды обитания (температура, pH и др.), наличие токсического загрязнения. Наиболее существенным фактором является смена видового состава. Как известно, у водорослей различных отделов, кроме основного пигмента, хлорофилла а, в фотосинтезе участвуют и другие пигменты - сборщики энергии: хлорофиллы b и c, фикобилины, каратиноиды, имеющие иные максимумы поглощения. В связи с этим определения коэффициентов регрессионных зависимостей (калибровка флюориметров) должны производиться для каждого изучаемого водоема и сезона. Однако, изменения видового состава фитопланктона, существенно влияющее на удельный выход флюоресценции, могут происходить достаточно быстро, особенно в небольших водоемах, где доминанты меняются каждые три-пять дней. Данные факторы приводят к необходимости постоянной калибровки флюориметров, что снижает пространственно-временное разрешение при зондировании поля флюоресценции.
Перечисленные недостатки флюоресцентного метода были преодолены группой сотрудников Красноярского Госуниверситета под руководством В.М.Гольда. В 1983 г. ими был разработан способ дифференциальной флюоресцентной оценки концентрации хлорофилла по основным отделам водорослей. Для этого используются флюориметры с переменной длиной волны возбуждающего света, действующего на основные пигменты-сборщики энергии, характерные для каждой группы водорослей. В большинстве озер и водохранилищ доминируют водоросли трех отделов: 1) сине-зеленые (Cyanophyta), 2) диатомовые (Bacillariophyta) и 3) зеленые (Chlorophyta). Для этих отделов наиболее эффективными в возбуждении флюоресценции являются, соответственно, следующие области спектра: 1) зеленая, с максимумом на длине волны l = 540 нм, 2) сине-зеленая, l = 510 нм и 3) синяя, l = 410 нм.
При дифференциальном способе калибровка флюориметра сводится к определению удельных выходов флюоресценции хлорофилла, R, у чистых культур зеленых (R зел), диатомовых (R диат) и сине-зеленых (R с-з) водорослей при возбуждении тремя участками спектра. Возможна калибровка и непосредственно на водоеме, при явном доминировании одного из отделов. Полученные численные значения подставляются в систему из трех линейных уравнений:
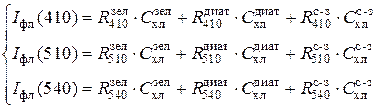 (1.1)
(1.1)
При рабочем использовании метода в пробе природного фитопланктона измеряются интенсивности флюоресценции на трех длинах волн (трех светофильтрах): I фл(410), I фл(510) и I фл(540). Затем, путем решения системы уравнений (1.1), находятся концентрации хлорофилла зеленых (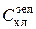 ), диатомовых (
), диатомовых (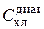 ) и сине-зеленых (
) и сине-зеленых (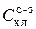 ) водорослей. Поскольку соотношения хлорофилл/биомасса относительно постоянны внутри каждого отдела, концентрация хлорофилла легко пересчитывается в биомассу. Таким образом, современные методы зондирования флюоресцентного поля позволяют получать информацию о биомассе (концентрации хлорофилла а) и видовом составе фитопланктона с очень высоким по сравнению со стандартными гидробиологическими методами пространственно-временным разрешением.
) водорослей. Поскольку соотношения хлорофилл/биомасса относительно постоянны внутри каждого отдела, концентрация хлорофилла легко пересчитывается в биомассу. Таким образом, современные методы зондирования флюоресцентного поля позволяют получать информацию о биомассе (концентрации хлорофилла а) и видовом составе фитопланктона с очень высоким по сравнению со стандартными гидробиологическими методами пространственно-временным разрешением.
Наряду с определением концентрации хлорофилла фитопланктона, изучение флюоресценции может дать информацию и об активности фотосинтетического аппарата водорослей, т.е., о потенциальной скорости фотосинтеза (первичной продукции). Для этого используются так называемые индукционные переходы флюоресценции. Если активную культуру микроводорослей (или лист высших растений) выдержать некоторое время в темноте, а затем подвергнуть освещению, флюоресценция вначале резко увеличится до максимального значения, Iмакс, а затем спадет до некоторого стационарного уровня, Iст. Индукционные переходы флюоресценции наблюдаются только в живых культурах, при наличии функционирующего фотосинтетического аппарата и могут служить мерой активности фотосинтеза.
На практике измеряют величину максимального уровня флюоресценции, искусственно блокируя электрон-транспортную цепь. Чаще всего блокировка осуществляется путем добавления в пробу специального ингибитора, который останавливает транспорт электрона по ЭТЦ. Обычно в качестве ингибитора фотосинтетического транспорта электронов используются гербициды: диурон (номенклатурное название 3-(3,4-дихлорфенил)-1,1-диметилмочевина, английская аббревиатура DCMU) или симазин (2-хлор-4,6- бис -(этиламино)-1,3,5-триазин). Величина разности между максимальным уровнем флюоресценции при добавлении и игибитора и ее стационарным значением D I = I макс(ингиб) - I ст, является достаточно надежной мерой активности фотосинтетического аппарата. При полевых исследованиях потенциальную величину фотосинтеза (первичной продукции) фитопланктона удобно определять по соотношению D I / I ст.
Флюоресцентные зонды могут решать задачу визуализации пространственного распределения фитопланктона для прицельного отбора гидробиологических проб и уточнения картины пространственно-временного распределения фитопланктона. Однако данных об использовании флюориметров в режиме непрерывной регистрации во внутренних водоемах сравнительно мало. Гораздо чаще публикуются сведения о точечных измерениях. Но даже точечный режим эксплуатации флюоресцентных приборов позволяет существенно увеличить пространственно-временное разрешение гидробиологических съемок.
Тем не менее, вертикальное зондирование флюорецентного поля в пресных водах, подобно вертикальному зондированию биолюминесцентного поля в океане, открыло существенные возможности для оперативного мониторинга важнейших кинетических характеристик водных экос