 2020-08-05
2020-08-05 210
210По способу образования клубеньки бобовых растений подразделяются на два типа:
1-й тип — клубеньки возникают при делении клеток перицикла (корнеродного слоя), обычно расположенных против протоксилемы (первых по времени образования сосудов) — эндогенный тип образования клубеньков;
2-й тип — клубеньки происходят из коры корня в результате внедрения возбудителя в паренхимные клетки коры и эндодермы (внутреннего слоя первичной коры) — экзогенный тип образования клубеньков. В природе преобладает последний тип. Ткани центрального цилиндра корня принимают участие только в образовании сосудистой системы клубеньков как эндогенного, так и экзогенного типа. Несмотря на различные взгляды на природу возникновения клубеньков зкзо- и эндотипов, процесс развития их в основном одинаков. Однако ни тот, ни другой тип образования клубеньков ни в коем случае не следует отождествлять с процессом образования боковых корней, несмотря на то что существуют и отдельные черты сходства в их заложении. Так, формирование клубеньков и боковых корней происходит одновременно и к тому же в одной и той же зоне корня. В то же время ряд особенностей развития боковых корней и клубеньков подчеркивает глубокие различия в типе их формирования. Боковые корни возникают в перицикле. С первых же моментов развития они связаны с центральным цилиндром главного корня, от которого ответвляются центральные цилиндры боковых корней, и возникают они всегда против луча первичной древесины. Формирование клубенька, в отличие от бокового корня, возможно в любом месте. В самом начале формирования клубеньковой ткани сосудистой связи с центральным цилиндром корня нет, она возникает позднее. Сосуды обычно формируются по периферии клубенька. Они связаны с сосудами корня через зону трахеид и имеют собственную эндодерму.
Различие в характере возникновения клубеньков и боковых корней особенно четко наблюдается у сераделлы, поскольку коровая ткань главного корня этого растения — место возникновения первых клубеньков — состоит из относительно небольшого слоя клеток и клубеньки становятся видимыми очень быстро после инфицирования корня бактериями. Они образуют сначала выступы уплощенной формы на корне, что позволяет отличить их от конических выступов боковых корней. Клубеньки отличаются от боковых корней и рядом анатомических признаков: отсутствием центрального цилиндра, корневых чехликов и эпидермиса, наличием значительного слоя коры, покрывающей клубенек. Формирование клубеньков (бобовых растений происходит в период, когда корень имеет еще первичную структуру. Оно начинается с деления коровых клеток, расположенных на расстоянии 2—3 слоев от концов инфекционных нитей. Слои коры, пронизанные инфекционными нитями, остаются без изменения. В то же время у сераделлы деление коровых клеток возникает непосредственно под инфицированным корневым волоском, а у гороха деление клеток отмечается только в предпоследнем слое коры.
Деление с образованием радиальной структуры ткани продолжается до внутренних коровых клеток. Происходит оно без определенного направления, беспорядочно, и в результате этого возникает меристема (система образовательных тканей) клубенька, состоящая из мелких зернистых клеток. Разделившиеся клетки коры изменяются: ядра округляются и увеличиваются в размерах, особенно увеличиваются ядрышки. После митоза ядра расходятся и, не принимая первоначальной формы, вновь начинают делиться. Возникает вторичная меристема. Вскоре в эндодерме и перицикле появляются признаки начинающегося деления, которое в прежних внешних клетках происходит главным образом тангентальными перегородками. Это деление распространяется, наконец, на общий меристематический комплекс, мелкие клетки которого вытягиваются, вакуоли исчезают, ядро заполняет большую часть клетки. Образуется так называемый первичный клубенек, в плазме клеток которого клубеньковые бактерии отсутствуют, поскольку они на данной стадии еще находятся внутри инфекционных нитей. В то время как образуется первичный клубенек, инфекционные нити многократно разветвляются и могут проходить или между клетками— интерцеллюлярно, или сквозь клетки — интрацеллюлярно — и вносить бактерии. Межклеточные инфекционные нити вследствие активного размножения в них клубеньковых бактерий нередко приобретают причудливую форму — формируются в виде карманов (дивертикулов) или факелов. Процесс передвижения инфекционных нитей из клетки в клетку не совсем ясен. По-видимому, инфекционные нити, как полагает канадский микробиолог Д. Джордан (1963), блуждают в виде голых слизистых тяжей в межклеточных промежутках растительной ткани до тех пор, пока вследствие каких-то еще необъяснимых причин не начинают инвагинировать в цитоплазму примыкающих клеток. В некоторых случаях инвагинация инфекционной нити происходит в одну, в некоторых случаях — в каждую соседнюю клетку. По этим инвагинированным трубчатым полостям (дивертикулам) перетекает заключенное в слизь содержимое нити. 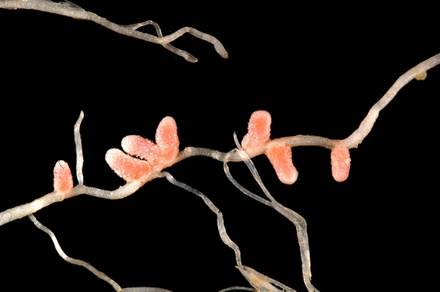 Наиболее активный рост инфекционных нитей происходит обычно вблизи ядра растительной клетки. Проникновение нити сопровождается перемещением ядра, которое продвигается к месту инфекции, увеличивается, меняет форму и дегенерирует. Подобная картина наблюдается при грибной инфекции, когда ядро нередко устремляется навстречу внедрившимся гифам, притягивается к повреждению как к месту наибольшей физиологической активности, вплотную придвигается к нити, разбухает и разрушается. Повидимому, это характерно для ответной реакции растения на инфекцию. У однолетних растений инфекционные нити возникают обычно в первый период инфицирования корня, у многолетних — в течение длительного периода развития. Бактерии могут высвобождаться из инфекционной нити в разное время и разными способами. Выход бактерий, как правило, весьма длительный процесс, особенно у многолетних растений. Обычно выход бактерий из инфекционной нити в цитоплазму растения-хозяина связывают с внутренним давлением, возникающим вследствие интенсивного размножения бактерий в нити и экскреции ими слизи. Иногда бактерии выскальзывают из нити группами, окруженными слизью инфекционной нити, в виде везикул (пузыревидных образований). Поскольку везикулы не имеют оболочек, выход из них бактерий очень прост. В клетки растений клубеньковые бактерии могут попадать и поодиночке из межклеточных пространств. Клубеньковые бактерии, вышедшие из инфекционной нити, продолжают размножаться в ткани хозяина. Размножение их в этот период происходит делением перетяжкой. Основная масса бактерий размножается в цитоплазме клетки, а не в инфекционной нити. Зараженные клетки дают начало будущей бактероидной ткани. Наполняющиеся быстро размножающимися клетками клубеньковых бактерий растительные клетки начинают усиленно делиться. В момент митотического деления зараженных клеток клубеньковые бактерии могут скапливаться на двух противоположных полюсах материнской клетки и пассивно попадать в дочерние клетки. Каждая из незаряженных клеток находится при этом под сильным стимулирующим воздействием клубеньковых бактерий и вследствие этого также делится. Благодаря такому энергично протекающему митотическому делению меристематических клеток осуществляется распространение клубеньковых бактерий в ткани клубенька и увеличение объема бактероидной области. Инфицированная ткань, состоящая из плотно лежащих и активно делящихся клеток, имеет сначала форму усеченного конуса. В дальнейшем вследствие постепенного роста этого конуса и одновременного деления и развития меристематических клеток ткань клубенька разрастается, утрачивая конусовидность. Таким образом, клубенек увеличивается сначала в результате радиального и тангентального деления коровых клеток, а затем за счет увеличения их размера и одновременного деления. После того как растительные клетки полностью заполнятся бактериями, митоз прекращается. Однако клетки продолжают увеличиваться в размере и часто сильно вытягиваются. Размер их в несколько раз больше, чем у неинфицированных растительных клеток, которые расположены между ними в бактероидной зоне клубенька.
Наиболее активный рост инфекционных нитей происходит обычно вблизи ядра растительной клетки. Проникновение нити сопровождается перемещением ядра, которое продвигается к месту инфекции, увеличивается, меняет форму и дегенерирует. Подобная картина наблюдается при грибной инфекции, когда ядро нередко устремляется навстречу внедрившимся гифам, притягивается к повреждению как к месту наибольшей физиологической активности, вплотную придвигается к нити, разбухает и разрушается. Повидимому, это характерно для ответной реакции растения на инфекцию. У однолетних растений инфекционные нити возникают обычно в первый период инфицирования корня, у многолетних — в течение длительного периода развития. Бактерии могут высвобождаться из инфекционной нити в разное время и разными способами. Выход бактерий, как правило, весьма длительный процесс, особенно у многолетних растений. Обычно выход бактерий из инфекционной нити в цитоплазму растения-хозяина связывают с внутренним давлением, возникающим вследствие интенсивного размножения бактерий в нити и экскреции ими слизи. Иногда бактерии выскальзывают из нити группами, окруженными слизью инфекционной нити, в виде везикул (пузыревидных образований). Поскольку везикулы не имеют оболочек, выход из них бактерий очень прост. В клетки растений клубеньковые бактерии могут попадать и поодиночке из межклеточных пространств. Клубеньковые бактерии, вышедшие из инфекционной нити, продолжают размножаться в ткани хозяина. Размножение их в этот период происходит делением перетяжкой. Основная масса бактерий размножается в цитоплазме клетки, а не в инфекционной нити. Зараженные клетки дают начало будущей бактероидной ткани. Наполняющиеся быстро размножающимися клетками клубеньковых бактерий растительные клетки начинают усиленно делиться. В момент митотического деления зараженных клеток клубеньковые бактерии могут скапливаться на двух противоположных полюсах материнской клетки и пассивно попадать в дочерние клетки. Каждая из незаряженных клеток находится при этом под сильным стимулирующим воздействием клубеньковых бактерий и вследствие этого также делится. Благодаря такому энергично протекающему митотическому делению меристематических клеток осуществляется распространение клубеньковых бактерий в ткани клубенька и увеличение объема бактероидной области. Инфицированная ткань, состоящая из плотно лежащих и активно делящихся клеток, имеет сначала форму усеченного конуса. В дальнейшем вследствие постепенного роста этого конуса и одновременного деления и развития меристематических клеток ткань клубенька разрастается, утрачивая конусовидность. Таким образом, клубенек увеличивается сначала в результате радиального и тангентального деления коровых клеток, а затем за счет увеличения их размера и одновременного деления. После того как растительные клетки полностью заполнятся бактериями, митоз прекращается. Однако клетки продолжают увеличиваться в размере и часто сильно вытягиваются. Размер их в несколько раз больше, чем у неинфицированных растительных клеток, которые расположены между ними в бактероидной зоне клубенька.
Большое влияние на жизнедеятельность клубеньковых бактерий и образование клубеньков оказывает реакция почвы. Для разных видов и даже штаммов клубеньковых бактерий значение рН среды обитания несколько различно. Так, например, клубеньковые бактерии клевера более устойчивы к низким значениям рН, чем клубеньковые бактерии люцерны. Очевидно, здесь также сказывается адаптация микроорганизмов к среде обитания. Клевер растет на более кислых почвах, чем люцерна. Реакция почвы как экологический фактор оказывает влияние на активность и вирулентность клубеньковых бактерий. Наиболее активные штаммы, как правило, легче выделить из почв с нейтральными значениями рН. В кислых почвах чаще встречаются неактивные и слабовирулентные штаммы. Кислая среда (рН 4,0— 4,5) оказывает непосредственное влияние и на растения, в частности нарушая синтетические процессы обмена веществ растений и нормальное развитие корневых волосков. В кислой среде у инокулированных растений резко сокращается срок функционирования бактероидной ткани, что ведет к снижению степени азотфиксации. В кислых почвах, как отмечает А. В. Петербургский, в почвенный раствор переходят соли алюминия и марганца, неблагоприятно действующие на развитие корневой системы растений и процесс азотоусвоения, а также снижается содержание усвояемых форм фосфора, кальция, молибдена и углекислоты. Неблагоприятную реакцию почвы лучше всего устраняет известкование. Размеры симбиотической азотфиксации определяются в значительной степени условиями питания растения-хозяина, а не клубеньковых бактерий. Клубеньковые бактерии как эндотрофные симбионты растений зависят в основном от растения при получении углеродсодержащих веществ и минеральных элементов питания. Для клубеньковых бактерий ткань хозяина представляет такую питательную среду, которая может удовлетворить даже самый требовательный штамм вследствие содержания в ткани всех типов питательных веществ. Тем не менее после внедрения клубеньковых бактерий в ткань растения-хозяина их развитие определяется не только внутренними процессами, но и в значительной степени зависит от действия внешних факторов, оказывающих влияние на весь ход инфекционного процесса. Содержание или отсутствие того или иного питательного вещества в окружающей среде может быть определяющим моментом для проявления симбиотической азотфиксации. Степень обеспеченности бобовых растений доступными формами минеральных соединений азота определяет эффективность симбиоза. На основании многочисленных лабораторных и вегетационных опытов известно, что чем больше азотсодержащих соединений в окружающей среде, тем с большим трудом внедряются бактерии в корень.
Большое значение в активации усвоения азота бобовыми растениями имеет фосфорное питание. При низком содержании фосфора в среде проникновение бактерий в корень происходит, но клубеньки при этом не образуются. Бобовым растениям присущи некоторые особенности в обмене фосфорсодержащих соединений. Семена бобовых отличаются повышенным содержанием фосфора. Запасной фосфор при прорастании семян используется не так, как У Других культур— сравнительно равномерно для формирования всех органов, а в большей степени сосредоточиваясь в корнях. Поэтому в ранние сроки развития бобовые растения, в отличие от злаковых, в большей степени удовлетворяют свои потребности в фосфоре за счет семядолей, а не запасов почвы. Чем крупнее семена, тем меньше бобовые растения зависят от фосфора почвы. Однако при симбиотическом способе существования потребность бобовых растений в фосфоре выше, чем при автотрофном. Поэтому при недостатке фосфора в среде у инокулированных растений ухудшается снабжение растений азотом.








