 2020-09-24
2020-09-24 692
692Тема: История э.
Эндокринология (от греч. endon — внутри, krino — выделять, logos — учение) — наука о железах внутренней секреции (специализированных органах, имеющих железистое строение) и выделяемых ими непосредственно в кровь гормонах (от греч. hormao — привожу в движение, возбуждаю). Термин «гормон» предложили У. Бейлисс и Э. Старлинг в 1902 г.
Эндокринология является общебиологической и общемедицинской дисциплиной. Общебиологическое значение эндокринологии состоит в первую очередь в том, что эта наука занимается изучением механизмов регуляции и интеграции функций. Эндокринная патология является заболеванием всего организма. Это обстоятельство, а также повсеместное учащение случаев эндокринной патологии определяют общемедицинское значение эндокринологии и требуют привлечения к борьбе с эндокринными заболеваниями врачей всех специальностей.
В развитии эндокринологии можно выделить 4 этапа: описательный; экспериментальный; выделение гормонов в чистом виде и расшифровка их химической структуры; синтез гормонов и получение их дериватов (производных).
Возникновение эндокринологии как науки относится к середине XIX в., когда в 1849 г. Бертольд впервые показал, что подсадка семенников в брюшную полость петухам после их кастрации предотвращает у них развитие посткастрационного синдрома. В том же году С. Е. Броун-Секар, удалив надпочечники у животных, доказал жизненную важность этих желез. В 1854 г. Шифф впервые отметил гибель животных после тиреоидэктомии. В 1884 г. пересадкой щитовидной железы тиреоидэктомированным животным он предотвратил их гибель и доказал этим роль щитовидной железы как органа внутренней секреции.
В 1855 г. Клод Бернар путем укола в дно IV желудочка мозга вызвал глюкозурию и гипергликемию и установил регулирующее воздействие нервной системы на функцию эндокринных желез. Клод Бернар ввел термин «внутренняя секреция».
В 1889 г. О. Минковский и И. Меринг экспериментально доказали связь между функцией поджелудочной железы и сахарным диабетом.
В 1889 г. Броун-Секар на заседании Парижского биологического общества сообщил об омолаживающих свойствах вытяжки из половых желез. Предположения Броун-Секара полностью не подтвердились, однако послужили поводом к применению органотерапевтических препаратов и гормонов для лечения больных. В 1901 г. Л. В. Соболев экспериментально доказал продукцию островковым аппаратом поджелудочной железы противодиабетического вещества инсулина и указал пути его получения.
Начало и середина XX в. были ознаменованы выделением из эндокринных органов ряда гормонов: адреналина (Такамине и Олдрич, к 1901), тироксина (Кендалл, 1915), инсулина (Бантинг и Бест, 1921), прогестерона (Бутенандт, 1934), адренокортикотропного гормона (Ли и Сайерс, 1943), трийодтиронина (Гросс и Лемблонд, 1950). В 1935 г. Дайзи синтезирован женский половой гормон эстрадиол, в 1954 г. был получен альдостерон — гормон клубочковой зоны коры надпочечников (Симпсон и Тайт), в 1963 г. обнаружен третий гормон щитовидной железы — тирокальцитонин (Копп). В 1957—1964 гг. Бергстрем и Ван-Дорп выделили в кристаллическом виде, установили химическую структуру и осуществили биосинтез ряда простагландинов. Из ткани опухоли островкового аппарата поджелудочной железы в 1966 г. впервые выделен проинсулин (Стейнер). Большим событием в эндокринологии было выделение из гипоталамуса рилизинг-факторов (рилизинг-гормонов), активирующих («либерины») или угнетающих («статины») продукцию тропных гормонов гипофиза. В 1962 г. из гипоталамуса выделен соматолиберин (соматотропин-рилизинг-фактор, Франц), в 1970 г.— тиролиберин (тиротропин-рилизинг-фактор, Шелли, 1968; Гиллемэн 1970), а в 1972 г. — соматостатин (соматотропин-рилизингингибирующий фактор, Гиллемэн).
В 1975—1978 гг. английские биохимики Д. Хьюз и Г. Костерлиц выделили из мозга свиньи, а затем и других животных эндогенные болеутоляющие вещества, относящиеся к группе пептидов, — энкефалины и эндорфины.
Одно из достижений современной эндокринологии — открытие иммунологических и радиоиммунологических методов определения уровня белковых гормонов в крови и моче. Благодаря использованию этих методов, обладающих высокой специфичностью, представилась возможность более точно по сравнению с биологическими тестами выявить изменения секреции, метаболизма и выделения белковых гормонов из организма. Большое внимание в настоящее время уделяется развитию нейроэндокринологии. Последняя изучает взаимодействие центральной нервной системы (ЦНС) и эндокринной системы в регуляции процессов жизнедеятельности. Исследования в области нейроэндокринологии и нейрофармакологии дают возможность углубленно изучать патогенез многих эндокринных заболеваний и способствуют внедрению более совершенных методов их диагностики.
Успехи современной эндокринологии, физиологии и морфологии позволяют более детально изучать механизмы регуляции различных функций организма.
Сравнительно недавно было установлено, что, помимо центральной (нервной) и эндокринной регуляции, существует еще и клеточная нейроэндокринная регуляция. В 1969 г. английский ученый А. Пирс сформулировал теорию, согласно которой в организме имеется функционально активная система, объединяющая эндокринные клетки, имеющие ряд общих свойств и общее происхождение с нервными клетками и продуцирующие белковые гормоны, а также биогенные амины, выполняющие функцию как гормона, так и нейромедиатора. Эта система получила название «АПУД-система» (от англ. amine content, precursor uptace decarboxylation — содержание аминов, поглощение предшественников и декарбоксилирование, определяющих принадлежность гормоно-образующей клетки к данной системе).
Успешно развиваются исследования желудочно-кишечных гормонов (эндокринология пищеварения). Установлено, что, помимо секретина, выделенного еще У. Бейлиссом и Э. Старлингом в 1902 г., нейроэндокринные клетки АПУД-системы (апудоциты) органов пищеварения синтезируют также следующие полипептидные гормоны: гастрин, холецистокинин-панкреозимин, мотилин, гастроингибирующий пептид (ГИП), вазоактивный интестинальный пептид (ВИП), субстанцию Р, бомбезин, нейротензин, энкефалин, соматостатин, глюкагон и энтероглюкагон, серотонин и мелатонин.
Широкое развитие в современной эндокринологии получило новое направление — иммунопатология. Изучение аутоиммунных процессов при эндокринных заболеваниях показало их важную роль в патогенезе диффузного токсического зоба, зоба Хасимото, первичного гипотиреоза, сахарного диабета, аддисоновой болезни и др. Сравнительно недавно была открыта система HLA (human leucocyte antigens — антигены лейкоцитов человека). Она определяет генетически детерминированный иммунологический ответ организма на различные антигены. Чрезвычайно перспективным является использование при изучении иммунопатологии моноклональных антител, методику получения которых разработали в 1975 г. Г. Кёлер и К. Мильштейн.
Тема: Современные представления о гормонах.
Гормон — специфические физиологические активные вещества, вырабатываемые специальными эндокринными органами или тканями, секретируемые в кровь или лимфу и оказывающие действие на функции и строение органов или тканей, вне места своего образования.
Ключевые признаки гормонов:
·активное вещество
·вырабатываемые специальными эндокринными органами
·выделяются в кровь или лимфу
·действие на клетку мишень
·дистантное действие
Не стоит путать гормоны с продуктами обмена веществ в организме, образующихся в результате жизнедеятельности различных клеток и обладающих определенной физиологической активностью — парагормоны.
Ех: углекислота, мочевина, свободные жирные кислоты, глюкоза
Гормоноиды — физиологически активные вещества, ктр не относятся согласно приведенного определения к гормонам.
Гормоноиды, парагормоны, разнородные по химической структуре биологически активные вещества, действующие на обмен веществ и многие физиологические процессы в организме. Образуются, как правило, не в железах внутренней секреции, как гормоны, а в др. тканях и органах. Биологическое действие Г. кратковременно, так как они разрушаются или связываются тканевыми белками. К Г. относят ацетилхолин, некоторые предшественники и продукты превращения адреналина, тканевые гормоны, или гистогормоны (гастрин, гепарин, секретин и др.), гистамин, серотонин и некоторые др.
Гормоноиды делятся на три группы:
1.«Клеточные» гормоны — физиологически активные вещества оказывающие свой эффект внутри тех клеток, в ктр они были образованы. Ех: простагландины
2.Тканевые г. - действие на ткань, выделившую г. Ех: циклические нуклеотиды
3.Медиаторы нервного возбуждения: ацетилхолин, серотонин, глютамат, гистамин, дофамин, норадреналин, гаммааминомасляная кислота (ГАМК)
Список некоторых известных веществ-нейромедиаторов:
· Адреналин
· Анандамид
· Аспартат
· АТФ
· Ацетилхолин
· Вазоактивный интестинальный пептид
· Дофамин
· ГАМК
· Гистамин
· Глицин
· Глутамат
· Глутаминовая кислота
· Норадреналин
· Окситоцин
· Серотонин
· Таурин
· Триптамин
· Эндоканнабиноиды
· N-ацетиласпартилглутамат
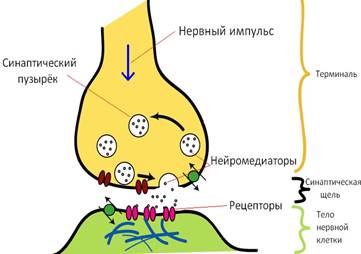
Действие медиаторов нервного возбуждения производится через передачу сигналов через синапсы.
Классификация гормонов
По эффекту ктр оказывают гормоны выделяют 4 группы:
1. Гормоны ктр оказывают метаболическое действие, т.е. вызывают изменение обмена веществ. Вообще, любой гормон изменяет обмен веществ. К 1 группе относятся только те гормоны, ктр вызывают изменение только определенного вида обмена веществ (белкового, жирового).
Ех: инсулин, глюкагон (углеводный обмен)
Кальцитонин (обмен Са)
Вазопрессин (обмен воды)
Ренин (водно-солевой обмен)
2. Г., ктр оказывают морфо-генетическое воздействие.
Заключается в стимуляции формообразовательных процессов, дифференциации органов и тканей, роста и метаморфоза. Ех.: половые гормоны.
3. Г., оказывающие кинетическое или пусковое действие, вызывающее определённую деятельность эффекторов.. Запускают какие-либо эффекты клеток. Без этих г. действие происходить не может. Например, «тропные» гормоны гипофиза необходимы для выработки и секреции гормонов щитовидной железы, половых желёз и некоторых гормонов коры надпочечников.
Ех.: рилизинг факторы гипоталамуса, тропные гормоны гипофиза.
Рилизинг-гормоны, или иначе рилизинг-факторы, либерины, релины — класс пептидных гормонов гипоталамуса, общим свойством которых является реализация их эффектов через стимуляцию синтеза и секреции в кровь тех или иных тропных гормонов передней доли гипофиза.
К известным рилизинг-гормонам относятся:
* кортикотропин-рилизинг-гормон
* соматотропин-рилизинг-гормон
* тиреотропин-рилизинг-гормон
* гонадотропин-рилизинг-гормон
Тропные гормоны, или тропины — подкласс гормонов передней доли гипофиза, реализующих своё физиологическое действие путём стимуляции синтеза и секреции гормонов периферических эндокринных желёз либо путём специфического «тропного» воздействия на определённые органы и ткани. Тропным гормоном регулируется активность эндокринных клеток пучковой зоны коры надпочечников, фолликуллов щитовидной железы, клубочковой зоны коры надпочечников, и НЕ регулируется активность околощитовидной железы.
К тропным гормонам относятся:
* адренокортикотропный гормон
* тиреотропный гормон
* фолликулостимулирующий гормон
* лютеинизирующий гормон
* соматотропный гормон
* пролактин
* меланоцитстимулирующий гормон
* липотропный гормон
β-эндорфин и мет-энкефалин, также производимые передней долей гипофиза, не относят к тропным гормонам.
4. - корригирующее, изменяющее интенсивность функций всего организма или его органов, которые могут совершаться на определённом уровне и без наличия гормонов.
Корригирующее действие гормонов состоит в изменении (усилении или ослаблении) тех физиологических процессов, которые могут осуществляться в организме и при отсутствии этих гормонов.
Адреналин, например, учащает ритм и увеличивает силу сокращений сердца, тормозит моторику желудочно-кишечного тракта, повышает тону мускулатуры многих кровеносных сосудов.
Классификация г. по химической структуре
|

 I. Белковые
I. Белковые
|
 |
|
 ТТГ — тиреотропный г.
ТТГ — тиреотропный г.
|
АКТГ-адренокортикотропный г. Вазопрессин ЛГ — лютеинизирующий г.
СТГ-соматотропный г. (АДГ-антидиуретический г.)
ЛТГ-лактотропный г. Окситоцин
Миланоцитостимулирующий г.
Паратиреоидный г.
Тиреокальцитонин
Инсулин, Глюкагон
II. Гормоны аминокислотной группы:
1) г. производные аминокислоты тирозина: адреналин, норадреналин, меланин — окисление тирозина ферментом тирозиназой; трийодтиронин, тироксин — йодированные производные тирозина, обр-зя в щитовидной железе.
2) г. производные аминокислоты триптофана: мелатонин (эпифиз)
III. Стероидные г.:
1) имеющие липидную природу
2) коры надпочечников: гидрокортизон, кортикостерон, кортизон, альдостерон; половые гормоны: эстрадиол, тестостерон, прогестерон
Механизм действия гормонов на клетки эфекторы
Наличие рецептора на клетке дает возможность г. действовать на эту клетку. Рецепторы могут располагаться на мембране, либо внутри клетки.
Клетка реагирует на тот или иной гормон только в том случае, если имеет к этому гормону рецептор. Специфический эффект гормона не связан ни с его синтезом, ни с транспортом, ни с активацией, ни с особенностями внутриклеточной реакции, специфика определяется лишь наличием рецептора гормона.
Локализация рецептора к гормону определяется способностью последнего проникать через клеточную мембрану.
По особенностями взаимодействия клеток и гормонов, последние могут быть разделены на 2 осоновных типа:
1 группа: белковые г. и катехоламины:
адреналин, норадреналин, дофамин
2 группа: стероидные и тироидные г.
Гормоны 1 группы плохо проникают внутрь клетки и действуют на ее поверхности, уже с самого начала требуют внутриклеточных медиаторов, опосредующих их эффекты.
Характерной стороной их действия является относительно быстрый эффект, обусловленный тем, что они активируют уже синтезированные в клетке ферменты и др. белки.
Гормоны 2 группы в свободном виде относительно легко проникают внутрь клетки и поэтому не требуют на первых этапах действия внутриклеточного посредника. Для их действия типична длительная и глубокая перестройка клеточного метаболизма.
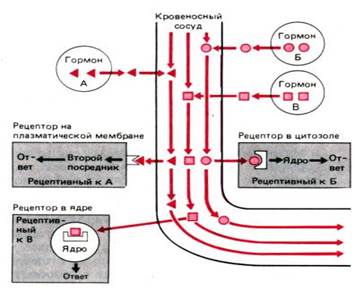 Механизмы действия гормонов. Три эндокринные клетки секретируют в межклеточное пространство в непосредственной близости от капилляров гормоны А, Б и В. Молекулы гормонов диффундируют в кровеносный сосуд и доставляются кровью к клеткам–мишеням, содержащим рецепторы этих гормонов. Гормон А взаимодействует с рецептором, находящимся на поверхности плазматической мембраны. Гормон–рецепторный комплекс стимулирует образование вторичного посредника, который индуцирует ответ клетки. Гормон Б взаимодействует с рецептором, находящимся в цитоплазме. Образующийся комплекс транслоцируется в ядро и воздействует на геном (например, изменяет синтез ядерной ДНК), что приводит к изменению синтеза белка. Гормон В взаимодействует с рецептором, локализованным в ядре, т.е. действует практически так же, как гормон Б, с той только разницей, что его рецептор исходно локализован в ядре
Механизмы действия гормонов. Три эндокринные клетки секретируют в межклеточное пространство в непосредственной близости от капилляров гормоны А, Б и В. Молекулы гормонов диффундируют в кровеносный сосуд и доставляются кровью к клеткам–мишеням, содержащим рецепторы этих гормонов. Гормон А взаимодействует с рецептором, находящимся на поверхности плазматической мембраны. Гормон–рецепторный комплекс стимулирует образование вторичного посредника, который индуцирует ответ клетки. Гормон Б взаимодействует с рецептором, находящимся в цитоплазме. Образующийся комплекс транслоцируется в ядро и воздействует на геном (например, изменяет синтез ядерной ДНК), что приводит к изменению синтеза белка. Гормон В взаимодействует с рецептором, локализованным в ядре, т.е. действует практически так же, как гормон Б, с той только разницей, что его рецептор исходно локализован в ядре
Механизм действия нестероидных гормонов
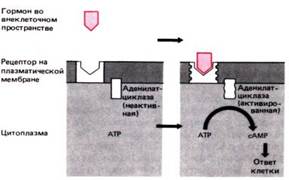 Механизм действия гормона, образующего комплекс с рецептором, локализованным на плазматической мембране. Достигнув рецептора и взаимодействуя с ним, гормон вызывает его трансформацию. Трансформация приводит к активации аденилатциклазы, также локализованной в плазматической мембране. Аденилатциклаза катализирует дефосфорилирование АТР с образованием сАМР. сАМР вызывает в клетке разнообразные эффекты, которые приводят в конечном счете к физиологическому ответу клетки. (АТР–аденозинтрифосфат, АТФ; сАМР–циклический аденозинмонофосфат, цАМФ)
Механизм действия гормона, образующего комплекс с рецептором, локализованным на плазматической мембране. Достигнув рецептора и взаимодействуя с ним, гормон вызывает его трансформацию. Трансформация приводит к активации аденилатциклазы, также локализованной в плазматической мембране. Аденилатциклаза катализирует дефосфорилирование АТР с образованием сАМР. сАМР вызывает в клетке разнообразные эффекты, которые приводят в конечном счете к физиологическому ответу клетки. (АТР–аденозинтрифосфат, АТФ; сАМР–циклический аденозинмонофосфат, цАМФ)
Рецептор локализован на поверхности мембраны и реакция клеток-эфекторов реализуется через циклазную систему, т.е. циклическая 3,5 АМФ формируется в 1 АТФ под влянием адениилатциклазы, а инактивируется фосфодиэстилазой.
Г. взаимодействует с рецептором, сигнал об этом взаимодействии передается на адениилатциклазу, локализованную на внутренней поверхности мембраны, или на фосфодиэстилазу.
При этом увеличивается или уменьшается образование циклического 3,5 АМФ, ктр в свою очередь активирует внутриклеточные ферменты.
Таким образом, именно 3,5 АМФ является тем внутриклеточным посредником, ктр обеспечивает передачу влияния г. на внутриклеточные системы. Характер же конечного ответа клетки определяется спецификой ферментного набора эффекторной клетки, а действие г. фактически оказывается толчком пускающим в ход реакцию предопределенную ферментативным профилем клетки.
Белковые гормоны, факторы роста, нейротрансмиттеры, катехоламины и простагландины относятся к группе гормонов, для которых рецепторы расположены на плазматических мембранах клеток.
Механизм действия стероидных гормонов
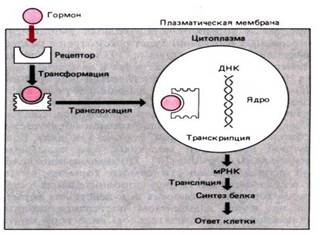 Взаимодействие гормона с внутриклеточным рецептором. Гормон диффундирует через плазматическую мембрану и взаимодействует с рецептором. Гормон–рецепторный комплекс переносится в ядро и действует на синтез ДНК, изменяя скорость транскрипции и количество информационной (матричной) РНК (мРНК). Увеличение или уменьшение количества мРНК влияет на синтез белка в процессе трансляции, что приводит к изменению функциональной активности клетки
Взаимодействие гормона с внутриклеточным рецептором. Гормон диффундирует через плазматическую мембрану и взаимодействует с рецептором. Гормон–рецепторный комплекс переносится в ядро и действует на синтез ДНК, изменяя скорость транскрипции и количество информационной (матричной) РНК (мРНК). Увеличение или уменьшение количества мРНК влияет на синтез белка в процессе трансляции, что приводит к изменению функциональной активности клетки
Циркулируя в крови стероиды присутствуют в виде свободных связанных форм. Белками плазмы связаны 98% половых гормонов, 90% кортизола, 50% альдостерона.
Гормон, находящийся в свободной форме, способен взаимодействовать с мембраной клетки и проходить через нее в цитоплазму, где связывается с цитоплазматическим рецептором, который отличается высокой специфичностью. Например, из гепатоцитов выделены рецепторные белки, связывающие только глюкокортикоидные гормоны или эстрогены. В настоящее время идентифицированы рецепторы к эстрадиолу, андрогенам, прогестерону, глюкокортикоидам, минералокортикоидам, витамину Д, тироидным гормонам, а также к ретиноивой кислоте и некоторым другим соединениям.
Поступление стероидов в клетку не зависит от мембранных барьеров или процессов транспорта.
Стероид в начале связывается в цитоплазме с плазматическим рецептором, ктр подвергается активации и комплекс рецептор-гормон перемещается из цитоплазмы в ядро и уже здесь происходит связывание комплекса ядерными акцепторными участками хроматина клеток-эфекторов.Это модулирует активность клеток специфических генов, ответственных за синтез определенных матричных РНК.
Механизм действия стероидных (жирорастворимых) гормонов
· Проникновение стероида (С) в клетку
· Образование комплекса СР (стероид-рецептор). Все Р стероидных гормонов представляют собой глобулярные белки примерно одинакового размера, с очень высоким сродством связывающие гормоны
· Трансформация СР в форму, способную связываться ядерными акцепторами [СР] Любая клетка содержит всю генетическую информацию. Однако при специализации клетки большая часть ДНК лишается возможности быть матрицей для синтеза иРНК. Это достигается путем сворачивания вокруг белков гистонов, что ведет к препятствию транскрипции. В связи с этим генетический материал клетки можно разделить на ДНК 3-х видов:
1.транскрипционно неактивная
2.постоянно экспрессируемая
3.индуцируемая гормонами или другими сигнальными молекулами.
· Связывание [СР] с хроматиновым акцептором.
Следует отметить, что этот этап действия С полностью не изучен и имеет ряд спорных моментов. Считается что [СР] взаимодействует со специфическими участками ДНК так, что это дает возможность РНК-полимеразе вступить в контакт к определенным доменам ДНК.
Интересным является опыт, который показал, что период полужизни иРНК при стимуляции гормоном увеличивается. Это приводит к многим противоречиям: становится непонятно увеличение количества иРНК свидетельствует, о том что [СР] повышает скорость транскрипции или увеличивает период полужизни иРНК; в то же время увеличение полужизни иРНК объясняется наличием большого числа рибосом в гормон-стимулированной клетке, которые стабилизируют иРНК или другим действием [СР] неизвестным для нас на сегодняшний момент.
· Избирательная инициация транскрипции специфических иРНК; координированный синтез тРНК и рРНК
Можно полагать, что основной эффект [СР] состоит в разрыхлении конденсированного хроматина, что ведет к открыванию доступа к нему молекул РНК-полимеразы. Повышение количества иРНК приводит к увеличению синтеза тРНК и рРНК.
· Процессинг первичных РНК
· Транспорт мРНК в цитоплазму
· Синтез белка
· Посттрансляционная модификация белка
Стероиды вначале связываются специфическими белками мембраны клетки, которые транспортируют их к цитоплазматическому рецептору или, минуя его, непосредственно к рецепторам ядра. Цитоплазматический рецептор состоит из двух субъединиц. В ядре клетки субъединица А, взаимодействуя с ДНК, триггирует (запускает) процесс транскрипции, а субъединица B связывается с негистоновыми белками. Эффект действия стероидных гормонов проявляется не сразу, а спустя определенное время, которое необходимо для образования РНК и последующего синтеза специфического белка.
Механизм действия тироидных гормонов
К тироидным г. относятся тироксин Т4 и трийодтиранин Т3. Основным гормоном щитовидной железы явл-ся Т4 и немного Т3.
Однако в клетках-эфекторах Т4 деиодируется в Т3, т.к. именно Т3 это главная внутриклеточная форма г. Т4 малоактивный г. Превращение Т4 в Т3 происходит в плазматической мембране и эндоплазматическом ретикуле. В качестве истинных рецепторов Т3 рассматриваются ядерные участки ответственные за действие тироидных г.
Тироидные гормоны (тироксин-Т4 и трийодтиронин-Т3), как и стероидные, легко диффундируют через липидную клеточную мембрану и связываются внутриклеточными белками. По другим данным, тироидные гормоны взаимодействуют сначала с рецептором на плазматической мембране, где комплексируются с белками, образуя так называемый внутриклеточный пул тироидных гормонов. Биологическое действие в основном осуществляется Т3, в то время как Т4 дейодируется, превращаясь в Т3, который связывается с цитоплазматическим рецептором. Если стероидцитоплазматический комплекс транслоцируется в ядро клетки, то тироидцитоплазматический комплекс сначала диссоциирует (растворяется, распадается) и Т3 непосредственно связывается рецепторами ядра, обладающими к нему высокой аффинностью(lat. affinitas: родственность). Кроме того, высокоаффинные рецепторы к Т3 обнаруживаются и в митохондриях. Считается, что калоригенное действие (увеличение основного обмена, сопровождающееся повышением теплообразования) тироидных гормонов осуществляется в митохондриях посредством генерации новой АТФ, для образования которой используется аденозиндифосфат (АДФ).
Тироидные гормоны регулируют синтез белка на уровне транскрипции и это их действие, обнаруживаемое через 12-24 часа, может быть блокировано введением ингибиторов синтеза РНК. Помимо внутриклеточного действия, тироидные гормоны стимулируют транспорт глюкозы и аминокислот через клеточную мембрану, непосредственно влияя на активность некоторых локализованных в ней ферментов.
Таким образом, специфическое действие гормона проявляется лишь после его комплексирования с соответствующим рецептором. В результате процессов по распознаванию, комплексированию и активированию рецептора последний генерирует ряд вторичных мессенджеров, которые вызывают последовательную цепь пострецепторных взаимодействий, заканчивающихся проявлением специфического биологического эффекта гормона.
Отсюда следует, что биологическое действие гормона зависит не только от его содержания в крови, но и от количества и функционального состояния рецепторов, а также от уровня функционирования пострецепторного механизма.
Синтез гормонов
1. Синтез тироидных горомонов.
Тироидные гормоны — г. щитовидной железы. Главными веществами в синтезе тироидных горомонов являются йод и тирозин.
Щитовидная железа способна захватывать йод из крови. Тирозин синтезируется организмом, а также может поступать с пищей, либо может распадываться из эндогенных продуктов.
Йод. Чтобы происходил синтез тиреоидных гормонов, суточное потребление иода должно составлять около 150 мкг. В кишечнике в процессе переваривания пищи йод отщепляется и всасывается в виде йодида и в этой форме циркулирует в крови в свободном состоянии.
В синтезе тироидных г. выделяют 6 этапов:
I. Биосинтез тиреоглобулина
II. Захват йодида
III. Органификация йодида
IV. Конденсация
V. Поглощение клетками и протеолиз (процесс ферментативного расщепления белков до пептидов и аминокислот) коллоида (некристаллизующиеся вещества, которые и в растворенном виде не проникают сквозь к.-л. Оболочки).
VI. Секреция
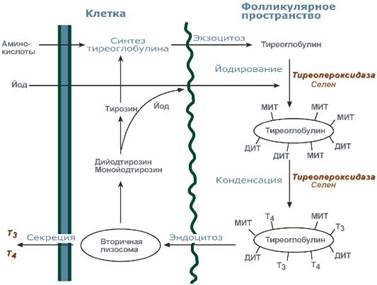
I. Тиреоглобулин синтезируется в фоликулярных клетках щитовидной железы и затем попадает в просвет фоликула щ.ж.
II. В просвет фоликула попадает йодид, ктр захватывается из крови этими же тиреоидными клетками
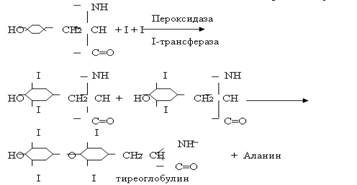
III. Под влиянием пероксидазы, цитохромоксидазы и флавин фермента — йодид окисляется в атомарный йод. И ковалентно связывается фенольными кольцами тирозиновых остатков, содержащихся в полипептидном каркасе тиреоглобулина.
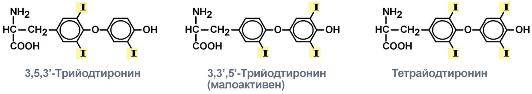
Связывание йода фенольным кольцом происходит только в 3-м, либо как в 3-м так и в 5-м положениях. Т.е. Присоединяется 1 или 2 молекулы йода.
Если присоединение происходит в 3 положении, то образуется монойодтирозин МИТ, если в 3 и 5, то дийодтирозин ДИТ.
Окисление йода может происходить и неферментным путем. Для этого необходимы ионы Сu и Fe.
Присоединение йода к тирозиновым остаткам называется органификацией. Соотношение МИТ и ДИТ в щитовидной железе составляет 1:3 или 2:3. Только 1/3 поглощенного йода используется для синтеза гормона, остальное удаляется с мочой.
IV. Этап конденсации йодтирозинов с образованием йодтиранинов
МИТ+ДИТ = Т3
ДИТ+ДИТ = Т4
Молекула тиреоглобулина не удаляется, конденсация происходит в молекуле.
В таком виде связанные с йодированным тиреоглобулином гормоны хранятся в фоликуле.
Комплекс йодированного тиреоглобулина называется коллоидом.???
Т.о. Тиреоглобулин составляет 10% от массы щ.ж., служит носителем и предшественником накапливания гормонов. Соотношение Т4 и Т3 7:1








