 2020-10-10
2020-10-10 472
472ПРЕВРАЩЕНИЯ ФОСФОРА
Фосфор – исключительно важный биогенный элемент. По значению в питании растений он занимает второе место после азота, хотя по содержанию в растениях он стоит среди других элементов только на 11 месте. Сосудистые растения поглощают фосфор в значительно меньших количествах, чем азот, кальций и калий. Тем не менее, его значение для растений очень велико, так как он входит в состав важнейших макромолекул клетки—некоторых белков, нуклеиновых и АТФ.
Фосфор составляет 0,12% массы земной коры, распределен в ней неравномерно и энергично мигрирует в биосфере. Он подвержен биогенному накоплению и миграции. Содержание Р2О5 в зональных почвах РФ выражается следующими показателями:
подзолистые 0,10
серые лесные и выщелоченные черноземы 0,13
обыкновенные и мощные черноземы 0,18
южные черноземы и каштановые почвы 0,15
сероземы 0,12
В 1 га пахотного слоя почвы содержится до 1—2 т валового фосфора, т. е. значительно больше, чем выносится с хорошим урожаем. Однако в некоторых южных районах фосфор находится в первом минимуме
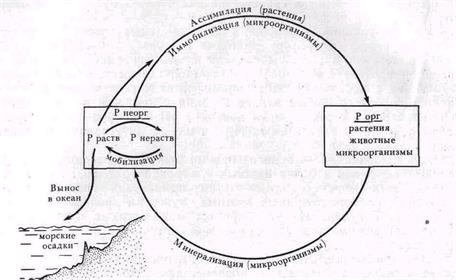
Рис 61 Превращения фосфора
и, как писал Д. Н. Прянишников, нужно добавить только один элемент — фосфор, чтобы оживить чернозем, истощенный многовековой культурой без удобрения. Дело в том, что фосфор в почве находится в труднодоступной для растений форме. Известно около 180 минералов фосфора, из которых наиболее распространены фосфаты кальция. Основным источником фосфора в почвах служат нерастворимые и слаборастворимые фосфорсодержащие минералы группы апатита, главным образом фторапатит.
Фосфор входит в состав многих органических соединений, которые содержатся в почве в живых телах, в остатках растений и животных, а также в гумусе. Фосфорорганические соединения составляют 10—50, а иногда до 80% всего запаса фосфора в почве. Коэффициент использования растениями фосфора из минеральных удобрений чрезвычайно низкий — всего 15—20% (ср. азота — 50% и калия 60—70%). У растений разных видов сильно различается реакция на обеспеченность почвы доступными соединениями фосфора. Очень сильное влияние на фосфорное питание растений оказывают микоризные грибы — симбионты корневых систем. Микосимбиотрофия распространена чрезвычайно широко. В настоящее время установлено наличие микоризы у 80% растений: у всех голосеменных и у 78% покрытосеменных. Она есть у растений разных жизненных форм — деревьев и кустарников, кустарничков и трав. Растения с микоризой встречаются во всех природных зонах, за исключением полярных пустынь и высокогорий.
Однако значение микроорганизмов в питании растений фосфором не сводится только к микосимбиотрофии. Свободноживущие микроорганизмы участвуют в процессах минерализации фосфорорганических соединений и способствуют переводу нерастворимых форм фосфора в растворимые. Эти процессы составляют основу превращения фосфора в природе (рис. 61).
МИНЕРАЛИЗАЦИЯ ФОСФОРОРГАНИЧЕСКИХ СОЕДИНЕНИЙ
Органические фосфорсодержащие соединения в почве входят в состав гумуса, торфа, навоза, растительных и животных остатков. Фосфор содержится в них, в противоположность азоту и сере, в окисленной форме, в виде остатка фосфорной кислоты. Наибольшая пропорция фосфорорганических соединений от общего запаса фосфора в почве содержится в черноземе (до 80%), а наименьшая — в сероземе (не более 10%). Большая их часть находится в форме фитина и фитатов, нуклеиновых кислот, фосфолипидов и гексозофосфатов. Живые клетки не способны поглощать большинство фосфорорганических соединений. Последние должны быть разложены до свободных фосфатных ионов, из которых затем вновь синтезируются внутриклеточно новые фосфорорганические вещества. Рассмотрим для примера разложение некоторых органических соединений, содержащих фосфор.
Фитин (соль инозитфосфорной кислоты) в кислых почвах закрепляется в виде солей Fe и Аl, а в щелочных и нейтральных — солей Са и Mg. Под действием микробных ферментов — фитаз — от фитина отщепляется 6 молекул Н3Р04. Лецитин и другие фосфолипиды — сложные эфиры глицерина и фосфорной кислоты — входят в состав цитоплазматических мембран. Расщепляются с участием внеклеточных микробных ферментов — фосфолипаз. Фосфорные эфиры сахаров гидролизуются неспецифическими фосфатазами. Фосфатазной активностью в той или иной степени обладают все почвенные микроорганизмы.
Нуклеиновые кислоты (РНК, ДНК) также содержат остатки фосфорной кислоты, которые освобождаются под действием микробных нуклеаз, выделяемых многими почвенными микроорганизмами.
МОБИЛИЗАЦИЯ НЕОРГАНИЧЕСКИХ СОЕДИНЕНИЙ ФОСФОРА
Фосфор в составе неорганических соединений входит в структуру первичных минералов или содержится в почве в виде нерастворимых солей Са, Fe и Аl. В качестве фосфорных удобрений применяют фосфориты и апатиты. Мобилизация из них фосфора происходит под действием кислот — органических и неорганических. Сильные неорганические кислоты образуют нитрификаторы (азотную) и тионовые бактерии (серную). Органические кислоты накапливаются в процессе анаэробных брожений и аэробных неполных окислении органических веществ грибами. Специфические органические кислоты продуцируют лишайники. Роль микоризных грибов в снабжении растений фосфором также определяется их способностью растворять фосфорсодержащие минералы путем выделения органических кислот.
Образующиеся под действием кислот микробного происхождения растворимые фосфаты иммобилизуются в клетках микроорганизмов, поглощаются растениями и частично вымываются в моря и океаны, где образуют осадки. Процесс этот однонаправленный, так как очень мало фосфора улетучивается в атмосферу или возвращается на сушу в виде гуано. Перевод нерастворимых фосфатов в доступную для растений форму — главное движущее звено в вовлечении фосфора в биологический цикл. Общий поток превращений фосфора представляет собой осадочный биогеохимический цикл, так как в океане образуются недоступные хранилища фосфора.
КРУГОВОРОТ СЕРЫ
Сера — один из необходимых для жизни биогенных элементов, входящих в состав некоторых аминокислот и растительных эфирных масел. В природе сера претерпевает химические и биологические превращения, переходя из неорганических соединений в органические и обратно. В виде неорганических соединений сера в почве бывает окисленная (сульфаты, политионаты), восстановленная (сульфиды и свободный H2S) и редко — молекулярная S2. При разложении остатков животных и растений освобождаются серусодержащие аминокислоты, где сера всегда находится в восстановленном состоянии.
Цикл превращений серы сходен с циклом азота: он включает окислительные и восстановительные звенья, а также звенья превращений без изменения валентности (рис. 62).
Ассимиляция сульфата растениями и микроорганизмами сопровождается восстановлением серы. Это так называемая ассимиляторная редукция серы, сходная с соответствующим процессом поглощения и восстановления нитратов. Процесс этот универсален для всех организмов. Биологическое закрепление растворимых сульфатов в микробных клетках носит также название иммобилизации серы. Минерализация серусодержащих органических соединений — процесс неспецифический. Он осуществляется микроорганизмами, которые обладают протеолитическими ферментами и разрушают белки. При аммонификации белков выделяется и аммиак и сероводород. В аэробных условиях может образовываться окисленная сера, и даже сульфаты, в анаэробных -- вместе с сероводородом выделяются летучие органические соединения типа меркаптанов.
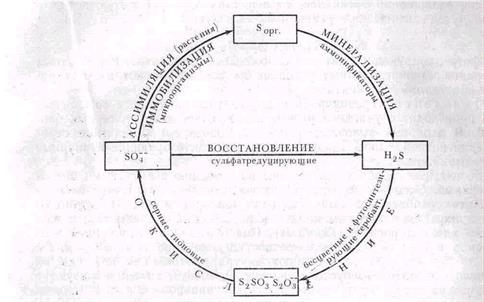
Рис 62 Круговорот серы
Сульфаты в анаэробных условиях восстанавливаются до сероводорода специфическими бактериями из группы облигатных анаэробов. Это узкоспециализированные микроорганизмы, которые используют сульфаты в качестве окислителя органических соединений в процессе анаэробного дыхания (ср. с денитрификацией). Они могут окислять и свободный водород, выступая как хемолитогетеротрофы: донором электрона для них служит Н2, а источником углерода — органические вещества, например, лактат. В последние годы выделены новые виды сульфатредуцирующих бактерий (Desulfonema limicola, Desulfosarcina variabilis), которые способны расти в автотрофных условиях, используя энергию окисления молекулярного водорода кислородом сульфатов для фиксации CO2. Возбудители процесса сульфатредукции относятся к разным родам анаэробных бактерий: Desulfovibrio — подвижные изогнутые палочки, не образующие спор, Desulfotomaculum — споровые палочки. Среди представителей первого рода есть галофилы, среди вторых — термофилы. Найдены сульфатредукторы и среди кокков (Desulfococcus, Desulfosarcina). Распространены сульфатредуцирующие бактерии в почвах с режимом, приводящим к длительному анаэробиозу, например в почвах затопляемых рисовых полей, а также в болотах, илах, лиманных грязях, в пластовых водах сопровождающих нефтяные месторождения. В подзолах мало сульфатов, и биогенным путем сульфиды в них не накапливаются. В щелочных и нейтральных почвах образуются нерастворимые сульфиды.
Накопление сульфида железа приводит к образованию черного ила. С жизнедеятельностью сульфатвосстанавливающих бактерий связывают процесс биогенного содонакопления. Провинции, где формируются содовые солончаки, приурочены к низменным местам, например, к поймам рек, понижениям вокруг болот. Если коренные породы в этих местах обогащены сульфатами, то в анаэробных условиях при затоплении и разложении органического вещества образуется H2S, который затем путем химических реакций преобразуется в Na2S и далее в соду:
Na2S + Н2О + СО2 > Nа2СОз + S
Сульфатредуцирующие бактерии способны к образованию карбонатных минералов. В определенных условиях их клетки бывают инкрустированы кристаллами кальцита.
Окисление сероводорода и других недоокисленных соединений серы происходит с участием разных специфических групп серных бактерий. В аэробных условиях окисление производят бесцветные серобактерии и тионовые, в анаэробных — фотосинтезирующие пурпурные и зеленые бактерии.
Бесцветные серобактерии сходны по строению с нитчатыми цианобактериями. Основное их отличие от последних — отсутствие пигмента. Нити серобактерий либо свободно плавают в воде (Beggiatoa), либо прикрепляются одним концом к неподвижным предметам, к растениям или водорослям (Thiothrix). Они обитают в кислородной зоне водоемов и производят окисление поступающего из анаэробной зоны H2S до S2, которую откладывают внутри клеток. Так как для их обильного развития требуется градиент О2 и H2S, то они образуют в толще воды пленку на границе аэробной и анаэробной зон. В Черном море, например, такая пленка, была обнаружена на глубине 200 м. Энергии окисления H2S до S2 оказывается достаточно для биологического связывания СОз. Поэтому серобактерии — хемолитоавтотрофы, использующие химическую энергию окисления неорганической серы в виде H2S для синтеза органических веществ из СО2. Впервые это явление исследовал С. Н. Виноградский в 1888 г. С этой работой связано открытие хемосинтеза у микроорганизмов.
Фотосинтезирующие серные бактерии, как и бесцветные нитчатые серобактерии, в основном обитатели водоемов, но в отличие от последних они анаэробы. Наличие в клетках пигментов бактериопурпурина или бактериохлорофилла обусловливает соответственно красную или зеленую окраску этих бактерий. Пурпурные бактерии (Chromatium) откладывают серу внутри клеток, а зеленые {Chlorobium} — вне клеток. В почвенных условиях они имеют небольшое значение, но могут быть активны на затопляемых участках почв рисовых полей. Фотосинтезирующие бактерии замыкают анаэробный цикл серы за счет окисления поступающего из анаэробной зоны H2S, образуемого сульфатредуцирующими бактериями.
Одноклеточные тионовые бактерии широко распространены в почвах. Впервые в чистую культуру тионовые бактерии выделил в 1904 г. М. Бейеринк, который дал им название Thiobacillus. Это название до сих пор сохраняется за одной из групп тионовых бактерий, хотя известно, что они не образуют спор. К настоящему времени среди тионовых бактерий описаны новые роды — Thiospirillum, Sulfolobus и Thiomicrospira. Эти бактерии в аэробных условиях окисляют H2S, сульфиды, тиосульфат, тетратионат, молекулярную серу и роданистые соединения до серной кислоты. Энергия окисления используется на синтез органических веществ из СОз через цикл Кальвина, как у всех автотрофных организмов. Акцептором электрона и водорода у них может выступать свободный кислород (Thiobacillus thiooxidans) или в анаэробных условиях — кислород нитратов (Th. denitrificans). Бактерии Th. ferrooxidans окисляют восстановленные соединения не только серы, но и железа. Среди тионовых бактерий есть уникальные виды, способные развиваться в очень кислых почвах с рН 3 и даже ниже (Th. Thiooxidans) или в щелочных (Th. thioparus).
Тионовые бактерии благодаря образованию сильной серной кислоты участвуют в природном сернокислотном выветривании, увеличивают подвижность нерастворимых фосфатов. Под действием этих бактерий происходит подкисление почв при добавлении серы к почвам со щелочной реакцией.
Тионовые бактерии используются и в практической деятельности для выщелачивания металлов из руд. Бактериальное выщелачивание металлов за счет окисления содержащихся в руде сульфидов позволяет вести добычу металла из бедного минерального сырья. Бактериальные методы выщелачивания обеспечивают один из этапов гидрометаллургии при минимальных энергозатратах.
Помимо биологического цикла серы в атмосфере Земли происходят превращения серусодержащих газов без участия живых организмов. Сера в виде сернистого газа образуется в результате сжигания нефти, а частично появляется в форме H2S и SO2 при извержении вулканов. Эти соединения окисляются в атмосфере, и образующаяся серная кислота возвращается на поверхность Земли и в почву. Вокруг промышленных районов таким путем происходит сильное подкисление почв вплоть до полной гибели растительности вблизи некоторых химических предприятий. Это одна из проблем охраны почв от техногенных загрязнений.
ПРЕВРАЩЕНИЯ КАЛИЯ
Калий — один из важнейших элементов, необходимых для растений. Он участвует в обменных процессах при синтезе аминокислот и белков, в реакциях фотосинтеза. Калий в значительной степени регулирует использование растениями азота. Между тем он относится к элементам, содержание которых в доступной форме в почве ниже потребности в них растений, и поэтому необходимо внесение калия в составе минеральных удобрений NPK
Усваиваемый калий составляет всего 1—2% от его общего количества в почве. Основной запас калия находится в минералах и в составе органоминеральных комплексов. Первичные минералы, содержащие калий, это слюды (биотит и мусковит) и полевые шпаты (ортоклазы и микроклины). Калий входит и во вторичные минералы: каолин, монтмориллонит, вермикулит. Освобождение калия из минералов происходит в процессе биологического выветривания. Процессы разложения минералов при взаимодействии с почвенными микроорганизмами и их метаболитами постоянно идут во всех почвах. В основе этих взаимодействий лежат разные механизмы: растворение сильными минеральными кислотами, образующимися при нитрификации, при окислении серы тионовыми бактериями; воздействие органических кислот — продуктов брожений и неполных окислении углеводов грибами; взаимодействие с внеклеточными аминокислотами, которые выделяются в среду многими почвенными микроорганизмами. С минералами взаимодействуют и продукты разложения микроорганизмами растительных остатков — полифенолы, таннины, полиурониды, флавоноиды, а также продукты микробного биосинтеза, например полисахариды. Некоторые из этих веществ действуют в растворе (кислоты), другие in situ на поверхности минералов (кислые гетерополисахариды). В опытах Н. Н. Сушкиной и И. П. Цурюпа с микроклином (8,7% К), биотитом (8,28% К) и бентонитом (1,89% К) в раствор вносили почвенные бактерии и через 100 дней обнаруживали освобождение из минералов более 50% калия. Рентгеноструктурный анализ подтвердил изменение крупных и мелких частиц алюмосиликатов под действием бактерий. Особенно активны в разрушении алюмосиликатов слизистые бактерии, выделяющие большое количество кислых полисахаридов. Среди них постоянно обнаруживается споровая палочка Bacillus circulans. Из неспоровых в превращении минералов участвуют представители рода Arthrobacter. В общем же это процессы неспецифические в которых участвуют разнообразные почвенные микроорганизмы.
ПРЕВРАЩЕНИЯ ЖЕЛЕЗА
Железо относится к элементам с переменной валентностью, и это обусловливает его разную подвижность в восстановительных и окислительных условиях. В первом случае оно активно мигрирует, во втором — инертно. Таким образом, основную роль в миграции железа в почве играет окислительно-восстановительная обстановка. В виде железоорганических соединений железо мигрирует легко, в виде FeS04 — слабо, а трехвалентное железо подвержено коллоидной миграции. Железо в почве содержится в следующих формах: в составе минералов (биотит, амфибол, пироксен); окисное нерастворимое (лимонит, гетит); растворимое — FеСОз (вивианит); псевдорастворимое — железо-гумусовые комплексы. В дерново-подзолистых почвах иллювиальные горизонты содержат 4—7% железа, а в красноземе его запас составляет 8-17%.
Участие микроорганизмов в превращениях железа в почве может быть прямым и косвенным. Рассмотрим несколько примеров.
1. Окисление железа в кислой среде происходит с участием специфических железобактерий, например Thiobacillus ferrooxidans. Процесс окисления Fe2+- Fe3+ идет за счет кислорода воздуха и с образованием энергии, которая используется в форме АТФ для фиксации: СО2 по типу хемосинтеза. Истинные железобактерии — хемолитоавтотрофы.
2. Окисление железа в нейтральной среде (в почвах гумидной зоны) - процесс побочный для осуществляющих его микроорганизмов и идет без использования ими энергии этого окисления. Как показала Г. А. Дубинина, он проводится микробами-гетеротрофами, которые удаляют этим путем образующуюся в их метаболических процессах перекись водорода. Эта функция у них проявляется только в специфических экологических условиях. К этой группе микроорганизмов относятся представители самых разных групп прокариот. Из них в почве распространены микоплазмы и представители рода Arthrobacter, а в воде — нитчатые, скользящие и цианобактерии.
Микоплазмы (класс Mollicutes) —сапрофиты, но в олиготрофных условиях способны паразитировать на других прокариотах или эукариотных организмах, например на грибах. Они окисляют железо и марганец, откладывая окислы на поверхности. Это мелкие полиморфные клетки, прорастающие нитями. К ним относятся Metallogenium symbioticum и Gallionella ferruginea.
Артробактерии — типичные почвенные бактерии со сложным циклом развития. К ним относятся многие бактерии, образующие отложения железа, которые были по-разному описаны на основании морфологических наблюдений их развития на стеклах обрастания или педоскопах: Siderocapsa, Siderobacter, Sideronema, Naumaniella, Ferribacterium.
3. Разложение железогумусовых комплексов в почве производят гетеротрофные микроорганизмы, осаждающие железо в виде конкреций и прослоек. Наиболее типичный для этого процесса организм - почвенная бактерия Seliberia stellata, описанная Т. В. Аристовской. Морфологически — это стебельковая бактерия, у которой стебельки закручены спирально, а отдельные клетки собираются в розетки.
4. Мобилизация железа из почвенных минералов происходит; за счет тех же механизмов, которые приводят к освобождению калия, т. е. благодаря образованию микроорганизмами слизей, фенольных соединений, щелочей. При этом образуются комплексные соединения, в которых железо мигрирует и аккумулируется в определенных горизонтах почв.
5. Восстановление железа при сопряженном окислении органического вещества или водорода происходит в анаэробных условиях с участием гетеротрофных микроорганизмов-полифагов (Bacillus или Clostridium) или анаэробных водородных бактерий из Pseudomonas, у которых Fe3+ - выступает в качестве акцептора электрона и может быть заменен молекулярным кислородом в аэробных условиях, или кислородом нитратов — в анаэробных. Восстановленное железо образует нерастворимый минерал — вивианит.
Таким образом, в превращениях железа в почве участвуют в основном микроорганизмы с неспецифическими функциями. Истинные железобактерии — группа облигатно-ацидофильных автотрофных бактерий, участвующие также и в превращениях серы.
ПРЕВРАЩЕНИЯ МАРГАНЦА
Марганец относится к группе необходимых для жизни микроэлементов. Его потребности покрываются микродозами, а в условиях его избытка наблюдается токсический эффект. В почве марганец находится в рассеянном состоянии в разных формах, в том числе и в виде металлорганических комплексов. Как и железо, он имеет переменную валентность и по-разному подвижен в зависимости от этого. Мигрирует марганец в почве главным образом в двухвалентной форме, в этой же форме он и усваивается растениями и микроорганизмами. В трех- и четырехвалентном состоянии марганец входит в состав железомарганцевых конкреций.
Бактерий, аналогичных железоокисляющим хемолитоавтотрофам в природе не обнаружено, хотя известны микроорганизмы, способные к аккумуляции и отложению марганца.
Превращения марганца в почве включают процессы его мобилизации из устойчивых природных соединений — минералов почвообразующих пород, образование и разрушение металлорганических комплексных соединений, биогенное окисление и аккумуляцию микроорганизмами, восстановление. Наиболее изученный микроорганизм, участвующий в окислении и аккумуляции марганца, — Metallogenium. В окислительных процессах, разрушении минералов и в разложении органоминеральных комплексных соединений с марганцем участвуют почвенные грибы, гетеротрофные бактерии, простекобактерии рода Pe domicrobium. Восстановление окислов марганца осуществляют факультативно-анаэробные микроорганизмы, которые понижают окислительно-восстановительный потенциал среды. Специфических восстановителей марганца типа денитрификаторов нет.
ПРЕВРАЩЕНИЯ АЛЮМИНИЯ
Алюминий — один из наиболее распространенных элементов на нашей планете и по содержанию в земной коре стоит на третьем месте после кислорода и кремния. В почве он находится в составе первичных и вторичных минералов, гидроокиси и алюмоорганических соединений, которые взаимно связаны (рис. 63). Мобилизация алюминия из кристаллических решеток алюмосиликатных минералов происходит при участии почвенных микроорганизмов, метаболиты которых либо образуют комплексные соединения непосредственно с алюминием либо выносят другие элементы из минералов, а алюминий, освобождаясь, связывается в металлорганические комплексы продуктами разложения растений или гумусовыми кислотами почв. В основе такого преобразования лежат реакции хелатизации. В форме металлорганических соединений алюминий приобретает подвижность и мигрирует в широком диапазоне рН. Эти процессы наиболее выражены в почвах зон влажного климата и связаны с подзоло- и латеритообразованием. В подзолистых почвах алюминий мигрирует в виде комплексных соединений с гумусовыми кислотами и накапливается в значительных количествах в иллювиальном горизонте в форме алюмоорганических комплексов с фульвокислотами.

Рис. 63. Превращения алюминия (по Т. В. Аристовской, 1980)
Алюмоорганические соединения не только образуются в самой почве, но и поступают в нее с растительными остатками в виде комплексов алюминия с органическими кислотами, аминокислотами, белками. Далее в зависимости от экологических условий алюмоорганические соединения в
Рис. 63. Превращения алюминия (по Т. В. Аристовской, 1980)
Алюмоорганические соединения не только образуются в самой почве, но и поступают в нее с растительными остатками в виде комплексов алюминия с органическими кислотами, аминокислотами, белками. Далее в зависимости от экологических условий алюмоорганические соединения в почве претерпевают различные превращения: 1) выносятся за пределы почвенного профиля; 2) минерализуются; 3) закрепляются в составе гумусовых веществ. Первые два процесса характерны для почв влажных субтропиков, третий — для почв подзолистой зоны.
Процессы минерализации алюмоорганических комплексных соединений связаны с жизнедеятельностью почвенных микроорганизмов. Экспериментальных работ в этой области пока очень мало. Т. В. Аристовская с сотрудниками показала, что в разложении этих веществ участвует комплекс грибов с организмами группы микоплазм — Metallogenium. В присутствии органоминеральных соединений алюминия или железа гифы грибов Penicillium sp. в симбиозе с Metallogenium покрываются отложениями гидроокисей соответствующего металла. Из этого можно заключить, что в определенной экологической обстановке микроорганизмы типа Metallogenium могут участвовать в накоплении железа и алюминия, а также марганца (см. выше) в почвенном профиле.
Накопление железа и алюминия в почвах тропической зоны при выносе кремния и оснований является, по-видимому, результатом активного поглощения полуторных окислов растениями, которые «перекачивают» эти элементы из нижних горизонтов в верхние (Родин, Базилевич, 1965).
ДРУГИЕ ЭЛЕМЕНТЫ
За счет включения разных механизмов почвенные микроорганизмы участвуют, по-видимому, в превращениях всех без исключения элементов, которые имеются в земной коре. Практически нет ни одного элемента, который тем или иным путем не подвергался бы воздействию микроорганизмов или их метаболитов. Одни элементы вовлекаются в биологический круговорот, входя в состав органических веществ, в процессе ассимиляции, другие окисляются, восстанавливаются или аккумулируются, третьи осаждаются или растворяются, извлекаются из минералов, подвергаются миграции, включаются в комплексы и т. д. и т. п.
Очень важны превращения, связанные с микроэлементами, которые необходимы микроорганизмам в очень малых дозах, но при этом роль их необычайно велика, так как они входят в состав ферментов и определяют их активность. Такова, например, роль молибдена, входящего в активный центр ферментов азотного цикла — нитрогеназы и нитратредуктазы; кобальта, ответственного за функции витамина B12; меди, которая включается в простетическую группу ферментов— оксидаз.
Известен ряд микроэлементов, которые не входят непосредственно в молекулы ферментов, но косвенно влияют на их активность и направление обмена веществ. К таковым относятся мышьяк, цинк, бор и др. Мышьяк, например, входит в состав всех живых клеток в микроколичествах. Его превращения в почве связаны с деятельностью микроорганизмов. Содержание мышьяка в почвах обычно невысокое — от 0,001 до 0,0001%. Оно наиболее высоко в черноземах и в органогенных горизонтах почв лесной зоны. Количество мышьяка увеличивается очень сильно за счет техногенных процессов. В его превращениях, аккумуляции и миграции участвуют грибы и бактерии. Окисление арсенита в арсенат — процесс биологический, о чем свидетельствует угнетение его азидом натрия, подавляющим дыхание. Среди бактерий, обусловливающих процесс окисления арсенитов, были описаны неспоровые грамотрицательные палочки из группы псевдомонад. Некоторые микроорганизмы ответственны за удаление мышьяка из почвы путем образования газообразных его соединений. Часть мышьяка фиксируется (иммобилизуется) клетками микроорганизмов за счет включения в обменные внутриклеточные реакции.
Бор в почвах в основном входит в состав органических соединений, из которых он освобождается микроорганизмами. Роль бора проявляется в образовании и функционировании клубеньков на бобовых растениях, так как он участвует в развитии сосудистой системы растений. Бор влияет на азотфиксацию клубеньковых бактерий, азотобактера и цианобактерий, а также стимулирует развитие многих аэробных и анаэробных бактерий и грибов. При малом содержании бора в почве или в условиях, затрудняющих его выведение из органических соединений, микроорганизмы выступают как конкуренты высших растений в отношении этого элемента, прочно удерживая его в своих клетках в составе органических веществ.
Для многих элементов известны только отдельные звенья превращений, связанных с деятельностью почвенных микроорганизмов. Есть сведения, например, о связывании цинка органическими соединениями в метаболизме грибов; о включении кобальта в молекулу B12 при синтезе этого витамина и др.
Микроорганизмы способны к аккумуляции в своих клетках элементов, которые содержатся в микроколичествах в почвах. Методом радиоавтографии показано, что микробные клетки накапливают естественно-радиоактивные элементы, такие как уран, торий, радий. На пленке после периода экспонирования микробных колоний, выращенных в присутствии указанных элементов, подсчитывают число треков — следов радиоактивного распада элементов, аккумулированных микроорганизмами.
Тионовая бактерия Thiobacillus ferrooxidans вызывает окисление сульфидов и косвенно влияет на миграцию таких элементов, как Cu, Zn, Ni и других, путем создания активного химического окислителя Fe2(SO4)3. Его взаимодействия с соединениями урана и ванадия изменяют валентное состояние этих элементов.
Известна роль микроорганизмов в процессах фракционирования стабильных изотопов некоторых элементов — углерода, азота, селена, лития. Примером может служить деятельность бактерий участвующих в превращениях серы. Первичная сера Земли представлена сульфидами. Изотопный состав серы в горных породах подвержен значительным колебаниям. За стандарт изотопного состава серы принимается сера метеоритов. Отклонения в соотношении легких (32S) и тяжелых (33S, 34S, 36S) изотопов серы есть результат последующего разделения атомов разного веса. Появление кислорода в газовой оболочке Земли привело к образованию сульфатов. При их восстановлении биологическим путем сульфатредуцирующие бактерии производят фракционирование изотопов серы: в H2S они переводят атомы легкой серы 32S, а в остаточном сульфате накапливается избыток тяжелых изотопа 34S. Таким образом, изотопный состав серы океанических сульфатов утяжелен на несколько процентов по сравнению с серой метеоритов, а сульфидные месторождения обогащены легким изотопом серы. По возрасту биогенных отложений серных руд делают попытки определить время появления кислорода на Земле, так как оно примерно совпадает с началом сульфатредукции.
В естественных биогеохимических провинциях, где почва обогащена молибденом, медью, ванадием, свинцом, бором, марганцем, почвенные микроорганизмы проявляют повышенную способность к связыванию этих элементов по сравнению с обедненными провинциями, что связано, возможно, с адаптационными явлениями в отношении этих геохимических факторов среды (Летунова, Ковальский, 1978).
Итак, вовлечение разных элементов в биологический круговорот идет разными путями и имеет разные механизмы. Углерод и азот вовлекаются в построение органических веществ и в основе их циклов лежат процессы синтеза и минерализации органических макромолекул, фиксация и возврат газообразных продуктов в атмосферу. Превращения фосфора и серы связаны с органическими веществами и разрушением минералов. В превращениях калия, железа, марганца, алюминия большую роль играют процессы разрушения и новообразования минералов, которые рассматриваются ниже. И превращения органических веществ, и разложение почвообразующих пород имеют две стороны: обеспечение растений элементами питания и роль в почвообразовании.
БИОЛОГИЧЕСКИЕ ПРОЦЕССЫ В ПОЧВООБРАЗОВАНИИ
Почвообразовательный процесс — это сложная совокупность явлений превращения и перемещения различных веществ в верхнем слое земной коры; взаимодействия малого биологического и большого геологического круговоротов веществ; баланса элементов, воды и тепла. Из разного сочетания этих явлений, которые в различной степени выражены в конкретных условиях, складывается определенный тип почвообразовательного процесса.
В каждом типе почвообразования можно выделить более общие процессы и частные, специфические. К общим относятся циклические процессы поступления и разложения органических веществ, биологической аккумуляции и выноса веществ из почвы, распада и синтеза вторичных минералов. Специфичны для каждого типа процессы гумусообразования и гумусонакопления, разложения первичных минералов и новообразования вторичных минералов. Среди частных почвообразовательных процессов выделяют макро- и микропроцессы. Первые охватывают всю почвенную толщу и ведут к образованию почвенного профиля, вторые сосредоточены в микрозонах, в отдельных очагах, что особенно характерно для некоторых микробиологических преобразований Процессы в пределах одного горизонта иногда называют мезопроцессами.
В последние годы в почвоведении развивается новое учение об элементарных почвенных процессах (ЭПП), основы которого были заложены трудами С. А. Захарова, С. С. Неуструева, Б. Б. Полынова. Под такими процессами понимают те, которые относятся только к почве. Они являются общими для всех типов, но их разное сочетание дает все разнообразие почв. И. П. Герасимов выделяет 13 таких элементарных почвенных процессов, другие исследователи – больше. Элементарными их назвали потому, что из них, как из кирпичиков, слагается весь почвообразовательный процесс. Эти процессы могут быть, в свою очередь, очень сложными. Некоторые из них имеют биологическую природу и в основе их лежит биохимический механизм.
Т.В. Аристовская (1980) предлагает по аналогии с ЭПП рассматривать и ЭПБП – элементарные почвенно-биологические процессы, относя к их числу следующие: 1 – разложение растительного опада; 2 – образование гумуса; 3 – разложение гумуса микроорганизмами; 4 – деструкция минералов почвообразующей породы почвенными микроорганизмами и их метаболитами; 5 – микробное минералообразование.
Кроме названных пяти процессов, которые обязательно проявляются во всех типах почв, есть еще ряд процессов характерных только для определенных типов почвообразования.








