2014-02-05
2014-02-05 5594
5594Экспрессия и репликация вирусного генома
Требования и ограничения
В ходе эволюции вирусов сложилось несколько стратегий, обеспечивающих: а) организацию вирусных генов и их кодирующую функцию, б) экспрессию вирусных генов, в) репликацию вирусных геномов и г) сборку и созревание вирусного потомства.
Прежде чем мы рассмотрим каждое из этих положений в деталях, стоит напомнить, что ключевым моментом в репликации вирусов является использование для синтеза вирусных белков хозяйских структур, синтезирующих белки клетки. Независимо от размеров, состава и организации своего генома вирус должен предоставить белоксинтезирующему аппарату эукариоти-ческой клетки информационную РНК, которую клетка должна распознать и транслировать. В этом отношении клетка навязывает вирусу два ограничения.
Во-первых, клетка синтезирует в ядре свою собственную мРНК путем транскрипции своей ДНК и последующего постранскрипционного процессинга транскрипта. Поэтому в клетке: а) ни в ядре, ни в цитоплазме нет фер* ментов, необходимых для транскрипции мРНК с вирусного РНК-генома, и б) в цитоплазме нет ферментов, способных транскрибировать вирусную ДНК. В связи с этим клеточную транскриптазу для синтеза вирусных мРНК могут использовать только вирусы, содержащие ДНК и способные проникать в ядро. Все другие вирусы вынуждены создавать собственные ферменты для синтеза мРНК.
Второе ограничение состоит в том, что синтезирующий аппарат эукариотических клеток приспособлен только для трансляции моноцистронных мРНК, так как он не распознает внутренних участков инициации в мРНК. В результате вирусы вынуждены синтезировать либо отдельные мРНК для каждого гена (функционально моноцистронная мРНК), либо мРНК, включающую несколько генов и кодирующую большой «полипротеин», который затем разрезается на индивидуальные белки.
Вирусные гены кодированы либо в РНК, либо в ДНК, которые могут быть либо одно-, либо двухцепочечными. Кроме того, геномы могут быть либо монолитными, когда все гены вируса содержатся в одной хромосоме, либо состоять из раздельных блоков, которые все вместе и составляют геном вируса. Во избежание ошибок мы обозначаем как «геномную» только нуклеиновую кислоту, которая заключена в вирионах.
Для удобства сначала лучше обсудить РНК-содержащие вирусы, фокусируя внимание прежде всего на функции геномной РНК.
Вирусы с одноцепочечной РНК можно подразделить на три группы. В первую группу входят пикорнавирусы и тогавирусы. Их геномы выполняют две функции (рис. 5.2 и 5.3). Во-первых, они функционируют как мРНК. Вирусы, геном которых может служить в качестве мРНК, принято называть вирусами с позитивным геномом. РНК пикорнавирусов после проникновения в клетку связывается с рибосомами и полностью транслируется. Затем продукт этой трансляции — полипротеин — расщепляется. Во-вторых, геномные РНК выполняют функцию матрицы для синтеза на ней комплементарных минус-цепей при участии полимеразы, появившейся в результате расщепления полипротеина. Образуется двуспиральная репликативная форма. На ее минус-цепях синтезируются новые плюс-цепи, которые могут использоваться в качестве а) мРНК, б) матриц для синтеза новых минус-цепей и в) составной части вирусных частиц потомства.
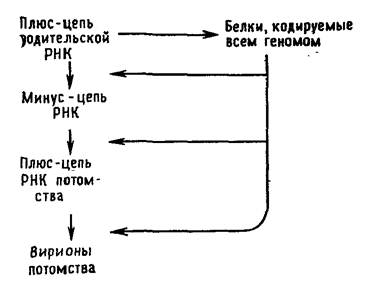
Размножение пикорнавирусов.
Тогавирусы и ряд других вирусов с позитивным геномом отличаются от пикорнавирусов в одном отношении: для трансляции в первом цикле синтеза белков доступна только часть их геномной РНК. Вероятной функцией образующихся при этом белков является способность транскрибировать геномную РНК. Они осуществляют синтез минус-цепи, которая в свою очередь служит матрицей для синтеза двух различных по размеру классов молекул плюс-РНК. В клетках, зараженных тогавирусами, РНК первого класса представлены небольшими молекулами мРНК, фланкирующими участок геномной РНК, который не транслировался в первом цикле. Образующиеся на них полипротеины расщепляются на белки, которые играют структурную роль в вирионах. Плюс-РНК второго класса состоит из полноразмерных цепей, которые упаковываются в вирионы.
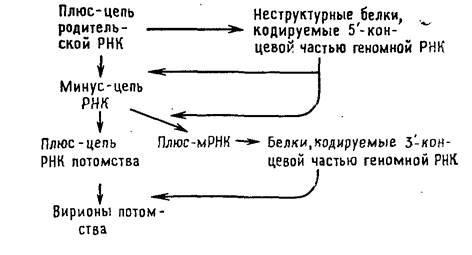
Размножение тогавирусов.
Главным в репликации вирусов с позитивным геномом является способность геномной РНК служить в качестве мРНК после заражения. Это имеет двоякие последствия. Во-первых, ферменты, ответственные за репликацию генома, синтезируются после заражения, и нет необходимости в их внесении в зараженную клетку вместе с вирионом. Вот почему «голая» РНК, экстрагированная из вирионов, инфекционна. Во-вторых, так как все позитивные геномы относятся к монолитным и все их гены сосредоточены в одной хромосоме, первичный продукт трансляции обеих РНК (как геномной, так и мРНК) обязательно представляет собой единый белок. Продукты трансляции пикорнавиру-сов и тогавирусов должны быть затем расщеплены на индивидуальные белки, которые и обнаруживаются в вирионе или в зараженной клетке.
РНК-содержащие вирусы с негативным геномом: ортомиксовирусы, парамиксовирусы, буньявирусы, аренавирусы и рабдовирусы.
Их геномная РНК выполняет две матричные функции: во-первых, для транскрипции и, во-вторых, для репликации. В связи с тем что для синтеза мРНК должен транскрибироваться вирусный геном, а в клетках соответствующие ферменты отсутствуют, все вирусы с негативным геномом содержат в вирионе кроме вирусного генома транскриптазу. Транскрипция вирусного генома — первое событие после проникновения вируса в клетку, в результате которого накапливаются функционально активные моноцистронные мРНК [позитивные, или плюс-цепи], кодирующие один белок. Репликацию начинают новосинтезированные вирусные белки, катализирующие образование полной плюс-цепи, которая служит матрицей для синтеза геномной минус-РНК (рис. 5.4).
Главное в репликации вирусов с негативным геномом заключается в том, что геномная РНК функционирует как матрица и для транскрипции, и для репликации. Отсюда следует, что, во-первых, вирус должен внести с собой в зараженную клетку транскриптазу; во-вторых, «голая» РНК, экстрагированная из вирионов, неинфекционна; в-третьих, синтезируемые мРНК имеют длину одного гена, они кодируют один белок — единичный полипептид. Присутствие сигналов сплайсинга в определенных участках может обеспечить формирование нескольких мРНК (каждая из которых кодирует особый белок) с одного и того же участка генома. Следовательно, плюс-транскрипт, функционирующий в качестве мРНК, отличается от плюс-РНК, служащей матрицей для вирусного потомства, хотя и первый, и вторая синтезируются на геномной РНК. Кроме РНК-транскриптазы в репликации вирусной РНК участвуют вирусиндуцированнные: репликаза 1 (обрзование репликативной формы) и репликаза 2 (синтез вирионных -нитей).
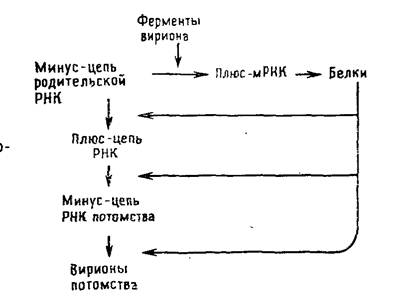
Размножение ортомиксовирусов и парамиксовирусов.
Ортомиксовирусы (вирусы гриппа А; В;С) генм представлен не одной а набором односпиральных РНК (грипп птиц и человека 8-7фрагментов). В структуре вириона:
– РНК зависимая РНК полимераза (синтез +нитей,
– две разновидности РНК полимераз
РНК транскриптаза (синтез +нитей в начале инфекции,
РНК репликаза (– нитей в конце инфекции)
Ретровирусы входят в третью группу РНК-содержащих вируссов.
Характерно, что геномы ретровирусов монолитны, но имеют диплоидную структуру, и обе цепи либо частично соединены водородными связями друг с другом, либо спарены неизвестным до настоящего времени образом. Единственная известная функция геномной РНК — матричная функция для синтеза вирусной ДНК. Поскольку эукариотические клетки не имеют для этого соответствующих ферментов, вирион кроме генома содержит еще и РНК-зависимую ДНК-полимеразу (обратную транскриптазу 1), а также смесь тРНК хозяина, одна из которых служит в качестве затравки. В цикле репродукции можно выделить следующие ключевые ступени: а) связывание комплекса тРНК —обратная транскриптаза с геномной РНК; б) синтез ДНК-копии, комплементарной по отношению к РНК, с переходом полимеразы с одной молекулы РНК-матрицы на другую, что приводит к образованию кольцевой одноцепочечной молекулы ДНК, связанной водородными связями с линейной геномной РНК; в) расщепление геномной РНК нуклеазой, атакующей только РНК в ДНК—РНК-гибридах (рибонуклеазои Н, также содержащейся в вирионе), и г) синтез комплементарной копии вирусной ДНК. Затем кольцевая двухцепочечная ДНК перемещается в ядро, где интегрирует с геномом хозяина, но последующая экспрессия вирусных генов не обязательна. Если экспрессия происходит, то интегрированная.вирусная ДНК транскрибируется транскриптазой клетки-хозяина. Продуктами транскрипции являются молекулы РНК, как равные по длине молекуле генома, так и более короткие мРНК-транскрипты нескольких соседних генов, которые транслируются с образованием полипротеинов. Полипротеины затем расщепляются на отдельные вирусные белки. В состав вирионов включаются только транскрипты, содержащие весь геном.
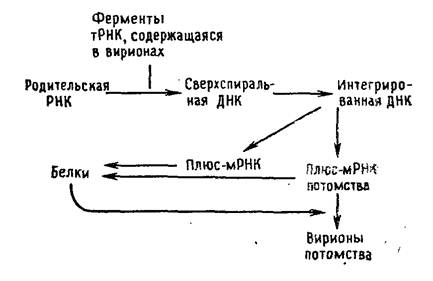
Размножение ретровирусов.