 2014-02-03
2014-02-03 750
750Для осуществления сокращения мышечного волокна требуется ряд условий:
1. Развитие потенциала действия на сарколемме.
2. Повышение внутриклеточной концентрации ионов кальция выше 0,1 мкМ.
3. Наличие в цитозоле молекул АТФ.
Потенциал действия (ПД) скелетного волокна имеет Na+,K+-ю природу (Рис.).
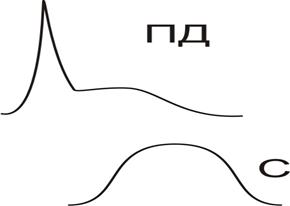
|
Его амплитуда достигает 120-130 мВ, овершут +30-+50 мВ, продолжительность 3-5 мс. Особенностью является длительная следовая деполяризация (около 10 мс), обусловленная аккумуляцией ионов K+ в Т-трубочках. Их разрушение при действии глицерина удаляет не только следовый потенциал, но и делает невозможным развитие сокращение на фоне ПД.
Повышение внутриклеточной концентрации ионов кальция происходит в результате его высвобождения из концевых цистерн СПР после развития ПД.
Взаимодействие электрических процессов на сарколемме (ПД) и непосредственно сократительных белков осуществляется с помощью процесса электромеханического сопряжения (ЭМС). В настоящее время существует несколько гипотез развития ЭМС:
1.Морфологическая, которая предполагает наличие структур, связывающих Т-систему и СПР.
2.Электрическая, которая предполагает перераспределение электрического поля с Т-системы на СПР.
3. Химическая, которая предполагает, что в процессе развития ПД внутрь волокна выделяется вещество-посредник запуска сокращения, например, инозитолтрифосфат.
После повышения внутриклеточного уровня выше критического (0,1 мкМ) ионы кальция соединяются с TrС, в результате чего весь тропониновый комплекс изменяет конформацию и снимается ингибиторное влияние TrI на молекулу актина. К открывающимся на молекуле актина участкам прикрепляются легкие цепи головки миозина. Формирующийся актин-миозиновый комплекс в присутствии АТФ осуществляет перемещение тонких и толстых миофиламентов относительно друг друга - теория скользящих нитей Хилла.
АТФ-азная активность миозина не подчиняется простой кинетике ферментативного катализа (Энгельгард В.А.,1939), так как протекает через ряд стадий, скорость которых зависит от структурных перестроек молекулы в присутствии актина и ионов кальция.
Лимитирующей стадией процесса становится распад фосфорилированной формы миозина в отсутствии актина (диссоциация актин-миозинового комплекса). Наоборот, после взаимодействия актина с миозином фосфорилированный миозин распадается очень быстро (скорость реакции увеличивается на несколько порядков). В результате распада молекула миозина становится свободной для очередного фосфорилирования, необходимого для перемещения нитей актина относительно миозина – собственно процесса сокращения.
Процесс расслабления начинается со снижения уровня внеклеточного кальция за счет работы системы активного транспорта СПР – Mg2+зависмойCa2+-АТФазы. Считается что этот процесс закачки внутрь СПР, в большей мере, происходит в его продольных трубочках.








