 2014-02-09
2014-02-09 756
756ЗАДАНИЕ К ПРИМЕРУ
Таблица 2
Таблица 2
Пример
I.Откроем задание к примеру (стр. 42) и построим интервальный вариационный ряд. Объем выборки n = 280; число интервалов k = 10.
1. Просматривая выборку, находим
х min = -3,483; х max = 5,717.
Размах выборки ∆ = 5,717 - (-3,483) = 9,2. Длина каждого интервала 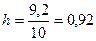 .
.
2. Запишем каждый из 10 интервалов (табл. 1).
3. Подсчитаем методом «конвертиков» абсолютные частоты ni, число выборочных данных, попадающих в каждый интервал, просматривая результаты наблюдений. Убедимся, что сумма всех частот равна 280: 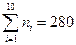 .
.
Шкала интервалов и группировка результатов наблюдений методом «конвертиков» приведены в табл. 1.
Таблица 1
| № | Интервал | Рабочее поле | n i |
| [-3,483; -2,563) | 
| ||
| [-2,563; -1,643) | 
| ||
| [-1,643; -0,723) | 
| ||
| [-0,723; 0,197) |     
| ||
| [-0,197; 1,117) | 
| ||
| [-0,117; 2,037) | 
| ||
| [2,037; 2,957) | 
| ||
| [2,957; 3,877) | 
| ||
| [3,877; 4,797) | 
| ||
| [4,797; 5,717] | 
|
4. Найдем центры каждого интервала по формуле
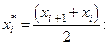
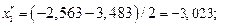
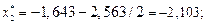
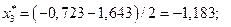
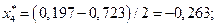
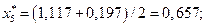
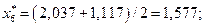
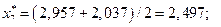
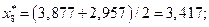
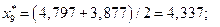

Данные вычислений занесем в табл. 2.
5. Вычислим для каждого интервала относительные частоты
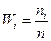
и плотность относительных частот 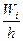 .
.
Убедимся, что сумма всех относительных частот равна единице:
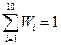 .
.
Данные вычислений занесем в табл. 2.
| № | Интервал | 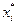
| n i | 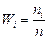
|
|
| [-3,483; -2,563) | -3,023 | 0,007 | 0,008 | ||
| [-2,563; -1,643) | -2,103 | 0,039 | 0,042 | ||
| [-1,643; -0,723) | -1,183 | 0,085 | 0,092 | ||
| [-0,723; 0,197) | -0,263 | 0,111 | 0,12 | ||
| [-0,197; 1,117) | 0,657 | 0,161 | 0,175 | ||
| [-0,117; 2,037) | 1,577 | 0,243 | 0,264 | ||
| [2,037; 2,957) | 2,497 | 0,179 | 0,195 | ||
| [2,957; 3,877) | 3,417 | 0,111 | 0,12 | ||
| [3,877; 4,797) | 4,337 | 0,050 | 0,054 | ||
| [4,797; 5,717] | 5,257 | 0,014 | 0,015 |
6. Построим гистограмму (рис. 1) и полигон относительных частот (рис. 2).
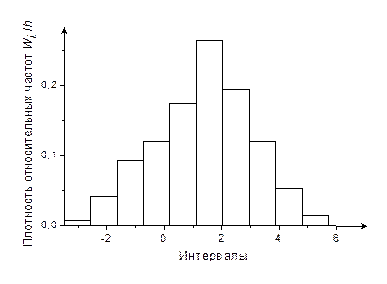
Рис. 1
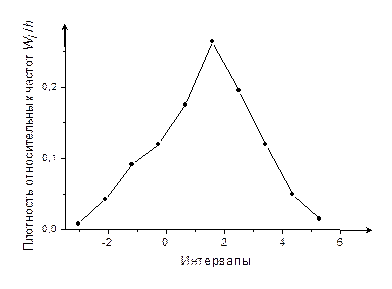
Рис. 2
II.Составим выборочную функцию распределения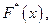 используя вариационный ряд относительных частот:
используя вариационный ряд относительных частот:
x ≤ -3,483 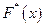 = 0;
= 0;
х Î (-3,483; -2,563] = W 1 = 0,007;
х Î (-2,563; -1,643] = W 1 + W 2 = 0,046;
х Î (-1,643; -0,723] = W 1 + W 2 + W 3 = 0,132;
х Î (-0,723; 0,197] = W 1 +W 2 + W 3 + W 4 = 0,243;
х Î (0,197; 1,117]) = W 1 +W 2 +W 3 + W 4 + W 5 = 0,404;
х Î (1,117; 2,037] = W 1 +W 2 +W3+W 4 +W 5 +W 6 = 0,646;
х Î (2,037; 2,957]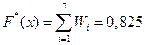 ;
;
х Î (2,957; 3,877] 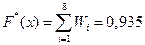 ;
;
х Î (3,877; 4,797] 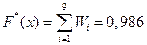 ;
;
х Î [4,797; 5,717]  ;
;
x > 5,717 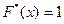 .
.
Запишем функцию:
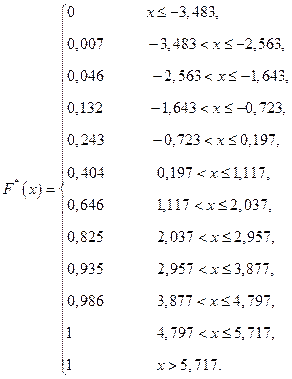
IV.Построим график функции (рис. 3).
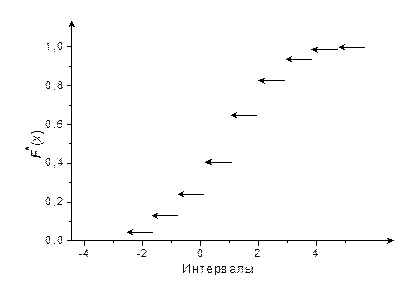
Рис. 3
III.Найдем числовые характеристики наблюдаемой случайной величины:
1. 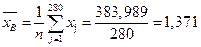 - выборочное среднее по всей выборке.
- выборочное среднее по всей выборке.
2. Среднее значение по сгруппированным данным
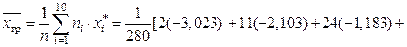
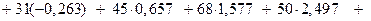
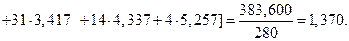
3. Вычислим дисперсию по сгруппированным данным:
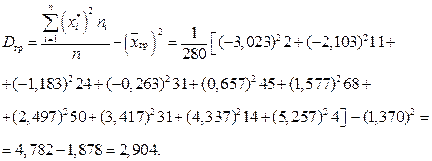
Среднее квадратическое отклонение по сгруппированным данным
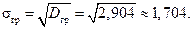
4. Относительная погрешность в вычислении среднего за счет замены выборки вариационным рядом
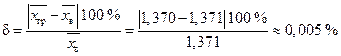 .
.
IV.Выдвигаем гипотезу о законе распределения генеральной совокупности.
Сравнивая полигон относительных частот (рис. 4) с видом графиков плотности распределения f (x) для основных непрерывных распределений (рис. 1-3), выдвигаем гипотезу, что случайная величина имеет нормальное распределение с параметрами m (математическое ожидание) и σ (среднее квадратическое отклонение).
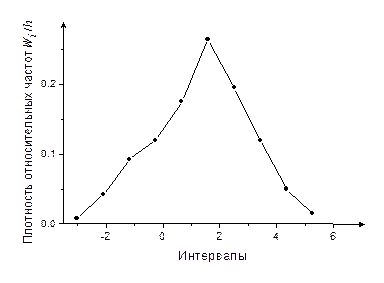
Рис. 4
V. Оценим параметры предполагаемого распределения и запишем его закон.
1. Оценим числовые характеристики исследуемой непрерывной случайной величины, используя оценки выборочных характеристик случайной величины Х:
математическое ожидание
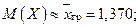
«исправленную дисперсию»
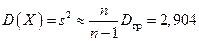 .
.
2. Согласно выдвинутой гипотезе о нормальном распределении случайной величины, запишем параметры данного распределения с учетом точечных оценок числовых характеристик предполагаемого распределения. Для нормального распределения – два параметра:
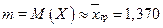 ,
,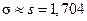 .
.
3. Запишем гипотетические функции плотности распределения f (x) и F (x), т. е. функции, соответствующие выдвинутой гипотезе о нормальном распределении:
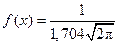
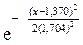 ;
;
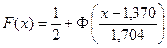 ,
,
где 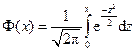 - функция Лапласа.
- функция Лапласа.
4. С помощью формул, соответствующих нормальному закону распределения, вычислим теоретические вероятности попадания случайного признака в каждый из 10 интервалов:
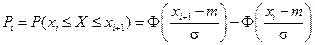 .
.
Значения функции Лапласа взяты с помощью таблицы (прил. 1).
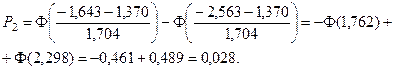
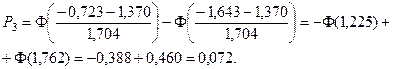
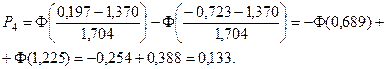
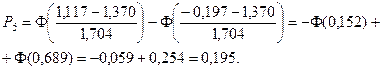
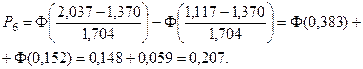
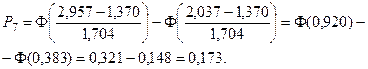
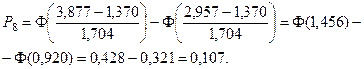
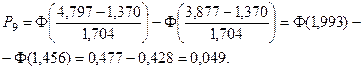
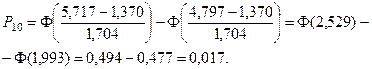
5. Найдем ожидаемые абсолютные частоты (теоретические) для данного закона распределения: 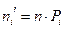 Полученные данные оформим в виде табл. 2.
Полученные данные оформим в виде табл. 2.
| Номер интервала |
| Рi | 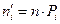
| 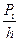
|
| -3,023 | 0,008 | 2,324 | 0.0087 | |
| -2,103 | 0,028 | 7,98 | 0.0304 | |
| -1,183 | 0,072 | 20,3 | 0.0783 | |
| -0,263 | 0,133 | 37,3 | 0.1446 | |
| 0,657 | 0,195 | 54,68 | 0.2120 | |
| 1,577 | 0,207 | 58,13 | 0.225 | |
| 2,497 | 0,173 | 48,49 | 0.1881 | |
| 3,417 | 0,107 | 29,87 | 0.1163 | |
| 4,337 | 0,049 | 13,664 | 0.0532 | |
| 5,257 | 0,017 | 4,872 | 0.0185 |
 Рис. 5.
Рис. 5.
6. Построим на одном графике (рис. 5) полигон относительных частот статистического распределения (кривая 1) (ломаная с вершинами в точках 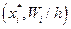 ) (рис. 4) и гипотетическую теоретическую плотность распределения, приближенно построенную по 10 точкам (кривая 2), (ломаная с вершинами в точках
) (рис. 4) и гипотетическую теоретическую плотность распределения, приближенно построенную по 10 точкам (кривая 2), (ломаная с вершинами в точках 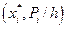 ).
).
Из графика видно, что теоретическая кривая распределения f (x), сохраняет в основном существенные особенности статистического распределения.
VI. Проведем статистическую проверку выдвинутой гипотезы.
1. Вычислим меру расхождения c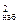 между теоретическим и статистическим распределениями по критерию Пирсона:
между теоретическим и статистическим распределениями по критерию Пирсона:
c2наб = 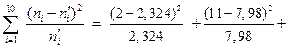
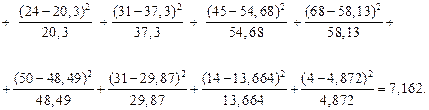
c2наб = 7,162.
2. Найдем по таблице (прил. 3) c2 критическое (c2кр). Уровень значимости примем α = 0,05; число степеней свободы r = k - - q - 1 = 10 - 2 - 1 = 7 (здесь k – число интервалов, q – число параметров предполагаемого распределения (q = 2). По входным параметрам таблицы (α = 0,05; r = 7) находим c2кр = 14,1.
3. Из сравнения cи c делаем вывод о правдоподобии гипотезы. Т. к. c2наб < c2кр, то экспериментальные данные не противоречат выдвинутой гипотезе, что случайная величина, представленная выборкой, имеет нормальный закон распределения.
делаем вывод о правдоподобии гипотезы. Т. к. c2наб < c2кр, то экспериментальные данные не противоречат выдвинутой гипотезе, что случайная величина, представленная выборкой, имеет нормальный закон распределения.
В результате выполненной работы на основе статистического материала получены закон распределения, эмпирическая функция распределения и числовые характеристики вариационного ряда. Вы научились подбирать плотность распределения непрерывной случайной величины, имея лишь выборку этой случайной величины. Кроме того, оценив параметры предполагаемого распределения, вы смогли проверить их соответствие наблюдаемым результатам. Именно с такой задачей сталкиваются на практике специалисты, применяющие методы математической статистики.
| 1,093 | -1,953 | -0,091 | -0,455 | 1,149 | 2,363 | 3,376 | -0,781 |
| 1,044 | 0,987 | 0,027 | 0,621 | -1,259 | 1,881 | 2,069 | 2,456 |
| 2,490 | 1,891 | 1,998 | 0,713 | 2,831 | 5,154 | -1,005 | 2,274 |
| -1,011 | 1,491 | -0,269 | 1,703 | 2,879 | 2,414 | 1,561 | 2,856 |
| -0,101 | 0,320 | 1,332 | 2,074 | 0,940 | 1,294 | 2,575 | 1,786 |
| 1,578 | 1,910 | 1,910 | 1,709 | 2,075 | -0,372 | 4,244 | -0,354 |
| 3,383 | -0,005 | 1,173 | 0,290 | 1,540 | -1,752 | -1,023 | 0,019 |
| -0,328 | 0,908 | 1,828 | 3,700 | 5,592 | 0,986 | 1,381 | 1,487 |
| -0,997 | -2,780 | 1,583 | 1,622 | 2,691 | 0,276 | 3,464 | 0,834 |
| 0,133 | 1,254 | -0,454 | 1,140 | 3,493 | 1,150 | 1,834 | 4,242 |
| 1,622 | 2,537 | 2,625 | 1,135 | 2,457 | 2,042 | 2,499 | 1,695 |
| -1,375 | 3,687 | -1,673 | 3,490 | 1,840 | -0,417 | 1,667 | 0,574 |
| 0,502 | -2,072 | 4,361 | 0,289 | 3,701 | -1,793 | 3,344 | 1,663 |
| 3,610 | 1,593 | 1,435 | 1,014 | 1,896 | 0,547 | 3,340 | 2,345 |
| 0,907 | 2,016 | 2,055 | 0,548 | 1,284 | 4,235 | 0,462 | 1,402 |
| 1,151 | 3,928 | 2,530 | 1,574 | 3,322 | 1,905 | -3,483 | 2,217 |
| 0,099 | 0,663 | 0,568 | -1,426 | 1,269 | 1,988 | -0,485 | 1,573 |
| 1,309 | 1,353 | 3,277 | 1,832 | 0,983 | -0,081 | 4,461 | 2,453 |
| 1,740 | 3,732 | 1,563 | 2,230 | 0,120 | -1,071 | 1,365 | 1,240 |
| 2,948 | -1,068 | 0,536 | 1,651 | 0,939 | 2,051 | 0,834 | -1,084 |
| 4,692 | -1,315 | 3,503 | 1,778 | 0,697 | -2,281 | -1,125 | 1,065 |
| 3,330 | 3,326 | -0,829 | 2,493 | -0,347 | 2,965 | 4,098 | 1,868 |
| -0,181 | 1,785 | 3,249 | 0,280 | -0,535 | 2,622 | -0,692 | 3,309 |
| 2,917 | 1,998 | 2,445 | -0,578 | 3,189 | 0,523 | 2,316 | -0,829 |
| 3,582 | -1,105 | 0,865 | 4,000 | 0,884 | 5,100 | -1,273 | 0,637 |
| 0,572 | 1,610 | -0,781 | 1,898 | 0,008 | 2,186 | 1,150 | -0,511 |
| 2,045 | 0,124 | 1,213 | 3,199 | -0,181 | 1,415 | 1,192 | 1,710 |
| 3,158 | -2,374 | 2,356 | 1,786 | 0,974 | 4,219 | -1,737 | 3,483 |
| 0,469 | -3,008 | 4,356 | 2,541 | 0,731 | -0,970 | -0,432 | -2,410 |
| 1,536 | 3,107 | -0,880 | 0,547 | 2,548 | 2,882 | -0,683 | 2,215 |
| 5,717 | 1,704 | 3,955 | 3,487 | 1,682 | 2,208 | -0,483 | 1,741 |
| -2,230 | 3,916 | 2,371 | 2,844 | -2,007 | 0,840 | -1,225 | 2,750 |
| 2,252 | -0,124 | 3,274 | 2,700 | 0,736 | -0,790 | 3,500 | 0,425 |
| -1,278 | 0,316 | 2,573 | 0,539 | -0,229 | 2,353 | 3,900 | 0,948 |
| 1,736 | 2,970 | 2,375 | 3,243 | 2,355 | 0,701 | 1,182 | 2,303 |
ПРИЛОЖЕНИЕ 1
Таблица значений функции 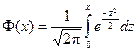
| х | |||||||||||
| 0,0 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 | 0,00000 | ||||||||||
| 1,0 1,1 1,2 1,3 1,4 1,5 1,6 1,7 1,8 1,9 | |||||||||||
| 2,0 2,1 2,2 2,3 2,4 2,5 2,6 2,7 2,8 2,9 | |||||||||||
| 3,0 3,5 4,0 4,5 5,0 | 0,49865 | 3,1 3,6 | 3,2 3,7 | 3,3 3,8 | 3,4 3,9 | ||||||
| - | - | - | - | - | - | - | - |
ПРИЛОЖЕНИЕ 2
Значения функции e - x
| х | e-x | r | x | e-x | r | x | e-x | r | x | e-x | r |
| 0,00 0,01 | 1,000 0,990 | 0,40 0,41 | 0,670 0,664 | 0,80 0,81 | 0,449 0,445 | 3,00 3,10 3,20 3,30 3,40 3,50 3,60 3,70 3,80 3,90 | 0,050 0,045 | ||||
| 0,10 | 0,905 | 0,50 | 0,606 | 0,90 | 0,407 | 4,00 4,10 4,20 4,30 4,40 4,50 4,60 4,70 7,80 7,90 | 0,0183 0,0091 | ||||
| 0,20 | 0,819 | 0,60 | 0,549 | 1,00 1,10 1,20 1,30 1,40 1,50 1,60 1,70 1,80 1,90 | 0,368 | 5,00 5,10 5,20 5,30 5,40 5,50 5,60 5,70 5,80 5,90 | 0,0067 | ||||
| 0,30 | 0,741 | 0,70 | 0,497 | 2,00 2,10 2,20 2,30 2,40 2,50 2,60 2,70 2,80 2,90 | 0,135 0,091 | 6,00 6,10 6,20 6,30 6,40 6,50 6.60 6,70 6,80 6,90 | 0,0025 | ||||
| 0,40 | 0,670 | - | 0,80 | 0,449 | - | 3,00 | 0,050 | - | 7,00 | 0,0009 | - |
| ПРИЛОЖЕНИЕ 3 | ||||||||||||||
| Значения c 2 в зависимости от r и a | ||||||||||||||
| r/a | 0,99 | 0,98 | 0,95 | 0,90 | 0,80 | 0,70 | 0,50 | 0,30 | 0,20 | 0,10 | 0,06 | 0,02 | 0,01 | 0,001 |
| 0,000 0,020 0,115 0,297 0,554 0,872 1,239 1,646 2,09 2,56 3,05 3,57 4,11 4,66 5,23 5,81 6,41 7,02 7,63 8,26 8,90 9,54 10,20 10,86 11,52 12,20 12,88 13,56 14,26 14,95 | 0,001 0,040 0,185 0,429 0,752 1,134 1,564 2,03 2,53 3,06 3,61 4,18 4,76 5,37 5,98 6,61 7,26 7,91 8,57 9,24 9,92 10,60 11,29 11,99 12,70 13,41 14,12 14,85 15,57 16,31 | 0,004 0,103 0,352 0,711 1,145 1,635 2,17 2,73 3,32 3,94 4,58 5,23 5,89 6,57 7,26 7,96 8,67 9,39 10,11 10,85 11,59 12,34 13,09 13,85 14,61 15,38 16,15 16,93 17,71 18,49 | 0,016 0,211 0,584 1,064 1,610 2,20 2,83 3,49 4,17 4,86 5,58 6,30 7,04 7,79 8,55 9,31 10,08 10,86 11,65 12,44 13,24 14,04 14,85 15,66 16,47 17,29 18,11 18,94 19,77 20,6 | 0,064 0,446 1,005 1,649 2,34 3,07 3,82 4,59 5,38 6,18 6,99 7,81 8,63 9,47 10,31 11,15 12,00 12,86 13,72 14,58 15,44 16,31 17,19 18,06 18,94 19,82 20,7 21,6 22,5 23,4 | 0,148 0,713 1,424 2,20 3,00 3,83 4,67 5,53 6,39 7,27 8,15 9,03 9,93 10,82 11,72 12,62 13,53 14,44 15,35 16,27 17,18 18,10 19,02 19,94 20,9 21,8 22,7 23,6 24,6 25,5 | 0,455 1,386 2,37 3,36 4,35 5,35 6,35 7,34 8,34 9,34 10,34 11,34 12,34 13,34 14,34 15,34 16,34 17,34 18,34 19,34 20,3 21,3 22,3 23,3 24,3 25,3 26,3 27,3 28,3 29,3 | 1,074 2,41 3,66 4,88 6,06 7,23 8,38 9,52 10,66 11,78 12,90 14,01 15,12 16,22 17,32 18,42 19,51 20,6 21,7 22,8 23,9 24,9 26,0 27,1 28,2 29,2 30,3 31,4 32,5 33,5 | 1,642 3,22 4,64 5,99 7,29 8,56 9,80 11,03 12,24 13,44 14,63 15,81 16,98 18,15 19,31 20,5 21,6 22,8 23,9 25,0 26,2 27,3 28,4 29,6 30,7 31,8 32,9 34,0 35,1 36,2 | 2,71 4,60 6,25 7,78 9,24 10,64 12,02 13,36 14,68 15,99 17,28 18,55 19,81 21,1 22,3 23,5 24,8 26,0 27,2 28,4 29,6 30,8 32,0 33,2 34,4 35,6 36,7 37,9 39,1 40,3 | 3,84 5,99 7,82 9,49 11,07 12,59 14,07 15,51 16,92 18,31 19,68 21,0 22,4 23,7 25,0 26,3 27,6 28,9 30,1 31,4 32,7 33,9 35,2 36,4 37,7 38,9 40,1 41,3 42,6 43,8 | 5,41 7,82 9,84 11,67 13,39 15,03 16,62 18,17 19,68 21,2 22,6 24,1 25,5 26,9 28,3 29,6 31,0 32,3 33,7 35,0 36,3 37,7 39,0 40,3 41,7 42,9 44,1 45,4 46,7 48,0 | 6,64 9,21 11,34 13,28 15,09 16,81 18,48 20,1 21,7 23,2 24,7 26,2 27,7 29,1 30,6 32,0 33,4 34,8 36,2 37,6 38,9 40,3 41,6 43,0 44,3 45,6 47,0 48,3 49,6 50,9 | 10,83 13,82 16,27 18,46 20,5 22,5 24,3 26,1 27,9 29,6 31,3 32,9 34,6 36,1 37,7 39,3 40,8 42,3 43,8 45,3 46,8 48,3 49,7 51,2 52,6 54,1 55,5 56,9 58,3 59,7 |
 | |||
 | |||
1. Микробиология — наука о возбудителях болезней. Зародилась как отрасль медицины.
В качестве самостоятельной науки, имеющей свои объекты и методы исследования, сформировалась во второй половине XIX в. благодаря работам Пастера, Коха, Эрлиха, Мечникова, Ру и др.
В настоящее время постоянно и интенсивно развивается, как и тесно связанные с ней биотехнология и генная инженерия.
В зависимости от решаемых задач микробиология делится на следующие отрасли:
• общая;
• промышленная;
• сельскохозяйственная; ветеринарная; санитарная;
• медицинская микробиология.
2. Предмет изучения медицинской микробиологии:
• микроорганизмы — представители нормальной микрофлоры тела человека и возбудители различных заболеваний человека;
• методы лабораторий диагностики, специфической профилактики и этиотропной терапии вызываемых ими заболеваний.
Вопрос 2. Классификация микроорганизмов
/. Понятие микроорганизмов
2. Систематика микроорганизмов
3. Прокариоты
4. Бактерии
1Микроорганизмы — это организмы, невидимые невооруженным глазом из-за их незначительных размеров.
Критерий размера — единственный, который их объединяет.
В остальном мир микроорганизмов еще более разнообразен, чем мир макроорганизмов.
2. Согласно современной систематике, микроорганизмы к 3 царствам:
• Vira — вирусы;
• Eucariotae — простейшие и грибы;
• Procariotae - истинные бактерии, риккетсии, хламидии, мико-плазмы, спирохеты, актиномицеты.
Так же как для растений и животных, для названия микроорганизмов применяется бинарная номенклатура, т. е. родовое и видовое название.
Если видовую принадлежность исследователям определить не удается и определена только принадлежность к роду, то употребляется термин species. Чаще всего это имеет место при идентификации микроорганизмов, имеющих нетрадиционные пищевые потребности или условия существования. Название рода обычно либо основано на морфологическом признаке соответствующего микроорганизма (Staphylococcus, Vibrio, Mycobacterium), либо является производным от фамилии автора, который открыл или изучил данный возбудитель (Neisseria, Shig-ella, Escherichia, Rickettsia, Gardnerella).
Видовое название часто связано с наименованием основного вызываемого этим микроорганизмом заболевания (Vibrio cholerae — холеры, Shigella dysenteriae — дизентерии, Mycobacterium tuberculosis — туберкулеза) или с основным местом обитания {Escherihia coli — кишечная палочка).
Кроме того, в русскоязычной медицинской литературе возможно использование соответствующего русифицированного названия бактерий (вместо Staphylococcus epidermidis — эпидер-мальный стафилококк; Staphylococcus aureus — золотистый стафилококк и т. д.).
3. Царство прокариот включает в себя отдел цианобактерий и отдел эубактерий, который, в свою очередь, подразделяется на порядки:
• собственно бактерии (отделы Gracilicutes, Firmicutes, Tenericutes, Mendosicutes);
• актиномицеты;
• спирохеты;
• риккетсии;
• хламидии.
Порядки подразделяются на группы.
Прокариоты отличаются от эукариот тем, что не имеют:
• морфологически оформленного ядра (нет ядерной мембраны и отсутствует ядрышко), его эквивалентом является нуклеоид, или генофор, представляющий собой замкнутую кольцевую двуни-тевую молекулу ДНК, прикрепленную в одной точке к цито-плазматической мембране; по аналогии с эукариотами эту молекулу называют хромосомной бактерией;
• сетчатого аппарата Гольджи;
• эндоплазматической сети;
• митохондрий.
Имеется также ряд признаков, или органелл, характерных для многих, но не для всех прокариот, которые позволяют отличать их от эукариотов:
• многочисленные инвагинации цитоплазматической мембраны, которые называются мезосомами, они связаны с нуклеоидом и участвуют в делении клетки, спорообразовании и дыхании бактериальной клетки;
• специфический компонент клеточной стенки — муреин, по химической структуре это пептидогликан (диаминопиеминовая кислота);
• плазмиды — автономно реплицирующиеся кольцевидные молекулы двунитевой ДНК с меньшей, чем хромосома бактерий, молекулярной массой. Они находятся наряду с нуклеоидом в цитоплазме, хотя могут быть и интегрированы в него, и несут наследственную информацию, не являющуюся жизненно необходимой для микробной клетки, но обеспечивающую ей те или иные селективные преимущества в окружающей среде.
Наиболее известны:
. F-плазмиды, обеспечивающие конъюгационный перенос
между бактериями;
. R-плазмиды — плазмиды лекарственной устойчивости, обеспечивающие циркуляцию среди бактерий генов, детерминирующих устойчивость к используемым для лечения различных заболеваний химиотерапевтическим средствам.
4. Бактерии — прокариотические, преимущественно одноклеточные микроорганизмы, которые могут также образовывать ассоциации (группы) сходных клеток, характеризующиеся клеточными, но не организменными сходствами.
Основные таксономические критерии, позволяющие отнести штаммы бактерий к той или иной группе:
• морфология микробных клеток (кокки, палочки, извитые);
• отношение к окраске по Граму — тинкториальные свойства (грамположительные и грамотрицательные);
• тип биологического окисления — аэробы, факультативные анаэробы, облигатные анаэробы;
• способность к спорообразованию.
Дальнейшая дифференциация групп на семейства, рода и виды, которые являются основной таксономической категорией, проводится на основании изучения биохимических свойств. Этот принцип положен в основу классификации бактерий, приведенной в специальных руководствах — определителях бактерий.
Вид является эволюционно сложившейся совокупностью особей, имеющих единый генотип, который в стандартных условиях проявляется сходными морфологическими, физиологическими, биохимическими признаками.
Для патогенных бактерий определение "вид" дополняется способностью вызывать определенные нозологические формы заболеваний.
Существует внутривидовая дифференцировка бактерий на
варианты:
• по биологическим свойствам - биовары или биотипы;
• биохимической активности— ферментовары;
• антигенному строению — серовары или серотжы;
• чувствительности к бактериофагам — фаговары или фаготипы;
• устойчивости к антибиотикам — резистентовары.
В микробиологии широко применяют специальные термины — культура, штамм, клон.
Культура — это видимая глазом совокупность бактерий на питательных средах.
Культуры могут быть чистыми (совокупность бактерий одного вида) и смешанными (совокупность бактерий 2 или более видов).
Штамм — это совокупность бактерий одного вида, выделенных из разных источников или из одного источника в разное время.
Штаммы могут различаться по некоторым признакам, не выходящим за пределы характеристики вида. Клон — это совокупность бактерий, являющихся потомством одной клетки.
Вопрос 3. Особенности морфологии микроорганизмов
1. Основные морфологические формы бактерий
2. Структура бактерий
3. Клеточная стенка
4. Цитоплазматическая мембрана
1 Среди основных морфологических форм бактерий различают: • шаровидные {кокковые);
• палочковидные.
Кокковые бактерии по характеру взаиморасположения делятся:
• на микрококки - отдельное изолированное расположение;
• диплококки - сцепленные попарно;
• тетракокки - сцепленные по четыре;
• стрептококки — сцепленные в цепочку;
• сарцины — сцепленные в пакеты по 8, 12, 16 и т. д.;
• стафилококки — сцепленные беспорядочно в виде виноградной грозди.
Палочковидные бактерии различаются:
• по форме:
• правильная — энтеробактерии, псевдомонады;
• неправильная — коринебактерии;
• размеру.
• мелкие - бруцеллы, бордетеллы;
. средние - бактероиды, кишечная палочка;. крупные — бациллы, клостридии;
• ф орме концов:
• обрубленные — бациллы;
• закругленные — сальмонеллы, псевдомонады;
• заостренные — фузобактерии; утолщенные — коринебактерии;
расположенные поодиночке;
диплобактерии и диплобациллы — сцепленные попарно;
• стрептобактерии и стрептобациллы — сцепленные в цепочку;
• извитые формы.
Извитые формы — по характеру и количеству завитков:
• вибрионы — слегка изогнутые палочки или неполные завитки;
• спириллы — один или несколько завитков;
• спирохеты, которые, в свою очередь, делятся:
• на лептоспиры (завитки с загнутыми крючкообразными концами — S-образная форма);
• боррелии (4—12 неправильных завитков);
• трепонемы (14—17 равномерных мелких завитков).
2. Структуру бактерий изучают в основном с помощью следующих методов:
• электронная микроскопия (техника ультратонких срезов);
• дифференциальное ультрацентрифугирование;
• цитохимия.
Структурные компоненты бактериальной клетки делятся на обязательные и необязательные.
Обязательные структурные компоненты:
• клеточная стенка;
• цитоплазматическая мембрана;
• цитоплазма с локализованными в ней рибосомами и ядерным аппаратом.
Необязательные структурные компоненты:
• капсула;
• микрокапсула;
• внеклеточная слизь;
• включения;
• жгутики;
• пили;
• споры.
3. Основу клеточной стенки у бактерий составляет пептидогликан муреин.
Функции клеточной стенки состоят в том, что она:
• является осмотическим барьером;
• определяет форму бактериальной клетки;
• защищает клетку от воздействий окружающей среды;
• несет разнообразные рецепторы, способствующие прикреплению фагов, колицинов, а также различных химических соединений,
• через клеточную стенку в клетку поступают питательные вещества и выделяются продукты обмена,
• в клеточной стенке локализован О-антиген и с ней связан эндотоксин (липид А) бактерий.
Существуют 2 типа строения клеточной стенки: %/ у бактерий первого типа пептидогликан муреин составляет до 90% массы клеточной стенки и образует многослойный (до 10 слоев) каркас, при этом он ковалентно связан с тейхоевыми кислотами. Такие бактерии при окраске по методу Грама прочно удерживают комплекс генцианового фиолетового и йода; они окрашиваются в сине-фиолетовый цвет и называются грамположителъными:
• у бактерий со вторым типом строения клеточной стенки поверх 2—3 слоев пептидогликана муреина располагается слой липополисахаридов. Эти бактерии при окраске по методу Грама неспособны прочно удерживать комплекс генцианового фиолетового и йода и соответственно обесцвечиваются спиртом, прокрашиваясь дополнительным красителем — фуксином — в розово-красный цвет. Они называются грамотрицательными.
В связи с различиями в строении клеточной стенки все бактерии делятся на 4 отдела:
• грациликуты — бактерии с тонкой клеточной стенкой, грамот-рицательные, к ним относятся различные извитые, палочковидные, кокковые формы бактерий, а также риккетсии и хла-мидии;
• фирмикуты — бактерии с толстой клеточной стенкой, грампо-ложительные, к ним относятся палочковидные, кокковые формы бактерий, а также актиномицеты, коринебактерии и микобактерии;
• тенерикуты — бактерии без ригидной клеточной стенки (микоплазмы);
• мендозикуты — архебактерии, отличающиеся дефектной клеточной стенкой, особенностями строения рибосом, мембран и рибосомальных РНК. Эта группа бактерий медицинского значения не имеет.
Из любой бактериальной клетки можно получить формы, полностью или частично лишенные клеточной стенки. Они называются соответственно протопластами и сферопластами и независимо от исходного морфологического типа бактерии из-за отсутствия клеточной стенки принимают шарообразную или грушевидную форму.
Кроме того, существуют L-формы бактерий, которые, в отличие от протопластов и сферопластов, способны к размножению, являясь вполне полноценными микробными клетками данного вида бактерий.
L-формы разных видов бактерий морфологически неразличимы. Независимо от формы исходной клетки (кокки, палочки, вибрионы) они представляют собой сферические образования разных размеров.
Имеются L-формы:
• стабильные — не реверсирующие в исходный морфотип;
• нестабильные — реверсирующие в исходный при устранении причины, вызвавшей их образование.
В процессе реверсии восстанавливается способность бактерий синтезировать пептидогликан муреин клеточной стенки. L-формы различных бактерий играют существенную роль в патогенезе многих хронических и рецидивирующих инфекционных заболеваний: бруцеллеза, туберкулеза, сифилиса, хронической гонореи и т. д.
4. К клеточной стенке бактерий примыкает цитоплазматическая мембрана, строение которой аналогично мембранам эукарио-тов — она состоит из двойного слоя липидов, главным образом фосфолипидов со встроенными поверхностными и интегральными белками).
Цитоплазматическая мембрана обеспечивает:
• селективную проницаемость и транспорт растворимых веществ в клетку;
• транспорт электронов и окислительное фосфорилирование;
• выделение гидролитических экзоферментов, биосинтез различных полимеров.
Кроме того, она ограничивает цитоплазму бактерий, которая представляет собой гранулярную структуру.
В цитоплазме локализованы рибосомы и бактериальный нуклеоид, в ней также могут находиться включения и плазмиды (вне-хромосомная ДНК).
Вопрос 4. Необязательные структурные компоненты бактериальной клетки
1. Споры
2. Жгутики
3. Ворсинки
4. Капсула
1. Споры бактерий представляют собой бактериальные клетки в состоянии анабиоза и образуются при неблагоприятных условиях внешней среды.
Располагаются внутри клетки терминально, субтерминально или центрально.
Спорообразуюише палочки называются бациллами. В проиессе спорообразования клетка почти полностью теряет воду, сморщивается, клеточная стенка уплотняется. Появляется новое вещество — дипиколинат кальция, которое образует комплексы с биополимерами клетки, устойчивые к действию температуры и ультрафиолетовых лучей.
В окружающей среде споры бактерий могут сохраняться годами, но при попадании в благоприятные условия спора впитывает влагу, комплексы распадаются, дипиколинат разрушается, и спора превращается в вегетативную клетку. Таким образом, спору следует рассматривать не как способ размножения, а только как форму существования бактериальной клетки в неблагоприятных условиях.
Преобразования идут по следующей схеме: 1 клетка — 1 спора — 1 клетка, т. е. увеличения количества бактериальных клеток не происходит.
Спорообразование характерно в основном для грамположитель-ных бактерий.
У грамотрицателъных бактерий эквивалентом спорообразования является переход в так называемое некультивируемое состояние. В такой форме они также длительно сохраняются в окружающей среде.
При использовании окраски по Граму споры не воспринимают красителей, поэтому на окрашенном фоне они бесцветны. Окрашиваются споры с помощью специальных методов окраски, например по Ожешко или Клейну.
2. Многие бактерии имеют жгутики, количество и расппппжение которых у разных родов неодинаково:
• монотрихии имеют только один жгутик (род Vibrio);
• лофотрихии — пучок жгутиков на одном полюсе клетки (род Pseudomonas);
• у амфитрихов жгутики (один или пучок) расположены на обоих полюсах клетки (род Spirillum);
• у перитрихов — по всей поверхности (род Escherichia, Salmonella). По своему строению жгутики представляют собой спирально закрученные нити, состоящие из специфического белка флагелли-на, который по своей структуре относится к сократительным белкам типа миозина.
При окраске по Граму жгутики не видны. Изучать подвижность бактерии можно как с помощью микроскопических методов (фазово-контрастная микроскопия препаратов "висячая" или "раздавленная" капля), так и посевом — уколом в полужидкий агар или специальную среду (среду Пешкова).
3. На поверхности ряда бактерий обнаружены белковые образования — ворсинки (фимбрии, пили).
Фимбрии отходят от поверхности клетки и состоят из белка, называемого пилином.
Различают более 60 видов ворсинок, из которых наиболее изучены следующие:
• F-pili — половые пили;
• common pili — пили, ответственные за адгезию.
4. Капсула бактерий — это утолщенный наружный слой клеточной стенки.
Капсулы могут быть построены из полисахаридов (пневмококк) или белков (возбудитель сибирской язвы).
Большинство бактерий, особенно патогенных, образует капсулу только в организме человека или животных. Однако существует род истинно капсулъных бактерий (Klebsiella), представители которого образуют капсулу и при культивировании на искусственных питательных средах.
Некоторые бактерии могут иметь микрокапсулу, выявляемую только при электронной микроскопии (эшерихии), или неявно выраженную способность к капсулообразованию — так называемую "нежную" капсулу (золотистые стафилококки, менингококки).
Основное предназначение капсул — зашита бактерий от фагоцитоза.
При окраске мазков по Граму истинно капсульные бактерии имеют характерное взаиморасположение (на расстоянии друг от друга). При световой микроскопии капсулы четко не видны, в связи с чем наличие капсул у бактерий выявляется с помощью специальных методов окраски, например по методу Гимзе. Для выявления капсул и бактерий, образующих их в организме, используют микроскопию мазков, приготовленных из патологического материала, или мазков-отпечатков из органов погибших животных.
Вопрос 5. Питание и особенности метаболизма бактерий
1. Химические компоненты бактериальной клетки
2. Питание бактерий
3. Метаболизм бактерий
1. Процесс, в ходе которого бактериальная клетка получает из окружающей среды компоненты, необходимые для построения ее биополимеров (органоидов), называется питанием.
Основными химическими компонентами бактериальной клетки являются органогены — кислород, водород, углерод, азот, фосфор. По химическому составу и характеру биополимеров (белки, полисахариды, нуклеиновые кислоты, липиды) прокариотические клетки не отличаются от эукариотических.
2. Бактериальные клетки не имеют специальных органов питания, т. е. являются голофитными.
Поступление питательных веществ в микробную клетку может происходить за счет:
• осмоса и диффузии по градиенту концентрации без затрат энергии;
• пассивного транспорта, который также осуществляется по градиенту концентрации с помощью белков-переносчиков, но без затрат клеткой энергии, и отличается от диффузии большей скоростью;
• активного транспорта, который идет против градиента концентрации с затратой энергии и возможным частичным расщеплением субстрата, осуществляется белками-переносчиками или ферментами — пермеазами.
По источникам углерода, необходимого для построения биополимеров, бактерии делятся на следующие группы:
• автотрофы — микроорганизмы, которые используют как единственный источник углерода углекислый газ и не нуждаются в сложных органических соединениях;
• гетеротрофы — микроорганизмы, которые используют в качестве источника углерода разнообразные органические углеро-досодержащие соединения (углеводы, углеводороды, аминокислоты, органические кислоты) как биологического, так и небиологического происхождения.
В зависимости от источника получения энергии микроорганизмы делятся:
• на фототрофные, способные использовать солнечную энергию,
• хемотрофные, получающие энергию за счет окислительно-восстановительных реакций.
В зависимости от природы доноров электронов:
• фототрофные литотрофы;
• хемотрофные литотрофы — использующие в качестве доноров электронов неорганические соединения;
• фото- и хемоорганотрофы — использующие только органические соединения. К последним принадлежит значительное большинство бактерий, в том числе патогенные для человека виды. По источникам азота:
• азотфиксирующие микроорганизмы — способны усваивать молекулярный азот атмосферы;
• микроорганизмы, ассимилирующие неорганический азот:
• солей аммония — аммонифицирующие;
• нитратов — нитратредуцирующие;
• нитритов — нитритредуцирующие.
Однако большинство патогенных для человека микроорганизмов способны ассимилировать только азот органических соединений.
Микроорганизмы, способные синтезировать все необходимые им органические соединения (углеводы, аминокислоты и др.) из указанных компонентов, называются прототрофами.
Микроорганизмы, неспособные синтезировать какое-либо из необходимых соединений и ассимилирующие их в готовом виде из окружающей среды или организма хозяина (человека, животного), называются ауксотрофами по этому соединению. Чаще всего ими являются патогенные или условно-патогенные для человека микроорганизмы.
3. Метаболизм (обмен веществ) бактерий представляет собой совокупность 2 взаимосвязанных противоположных процессов: катаболизма и анаболизма.
Катаболизм (диссимиляция) — распад веществ в процессе ферментативных реакций и накопление выделяемой при этом энергии в молекулах АТФ.
Анаболизм (ассимиляция) — синтез веществ с затратой энергии. Особенности метаболизма у бактерий состоят в том, что:
• его интенсивность имеет достаточно высокий уровень, что возможно обусловлено гораздо большим соотношением поверхности к единице массы, чем у многоклеточных;
• процессы диссимиляции преобладают над процессами ассимиляции;
• субстратный спектр потребляемых бактериями веществ очень широк — от углекислого газа, азота, нитритов, нитратов до органических соединений, включая антропогенные вещества — загрязнители окружающей среды (обеспечивая тем самым процессы ее самоочищения);
• бактерии имеют очень широкий набор различных ферментов — это также способствует высокой интенсивности метаболических процессов и широте субстратного спектра.
Ферменты бактерий по локализации делятся на 2 группы:
• экзоферменты — ферменты бактерий, выделяемые во внешнюю среду и действующие на субстрат вне клетки (протеазы, полисахариды, олигосахаридазы);
• эндоферменты — ферменты бактерий, действующие на субстраты внутри клетки (расщепляющие аминокислоты, моносахара, синтетазы).
Синтез ферментов генетически детерминирован, но регуляция их синтеза идет за счет прямой и обратной связи, т. е. для одних — репрессируется, а для других — индуцируется субстратом. Ферменты, синтез которых зависит от наличия соответствующего субстрата в среде (бета-галактозидаза, бета-лактамаза), называются индуцибельными.
Другая группа ферментов, синтез которых не зависит от наличия субстрата в среде, называется конститутивными (ферменты гликолиза). Их синтез имеет место всегда, и они всегда содержатся в микробных клетках в определенных концентрациях. Изучают метаболизм бактерий с помощью физико-химических и биохимических методов исследования в процессе культивирования бактерий в определенных условиях на специальных питательных средах, содержащих то или иное соединение в качестве субстрата для трансформации.
Такой подход позволяет судить об обмене веществ путем более, детального изучения процессов различных видов обмена (белков, углеводов) у микроорганизмов.
Вопрос 6. Особенности белкового и углеводного обмена у бактерий
1. Белковый обмен
2. Углеводный обмен
3. Типы биологического окисления у бактерий
1. Белковый обмен у бактерий:
- процесс синтеза собственных аминокислот и белков путем ассимиляции необходимых компонентов из внешней среды;
- внеклеточное расщепление белков под воздействием различных ферментов.
Если расщепление белков происходит в анаэробных условиях, то процесс называется гниением.
Если в аэробных условиях — тлением.
При наличии у бактерий протеаз белки расщепляются ими до промежуточных продуктов распада — пептонов. При наличии пептидаз пептоны расщепляются ими до аминокислот и продуктов их распада (аммиака, сероводорода, индола). Протеолитические (способность расщеплять белки) и пептоли-тические (способность расщеплять пептоны) свойства выражены далеко не у всех бактерий, поэтому их изучение в совокупности с другими ферментативными свойствами помогает идентифицировать бактерии.
2. Углеводный обмен у бактерий также носит двоякий характер — это процесс синтеза и распада углеводов.
Расщепление углеводов бактериями (сахаролитические свойства) в аэробных условиях с образованием углекислого газа и воды называется горением, а расщепление ими углеводов в анаэробных условиях — брожением.
В зависимости от характера конечных продуктов разложения углеводов в анаэробных условиях различают брожение:
• спиртовое;
• молочнокислое;
• пропионовокислое;
• муравьинокислое;
• маслянокислое;
• уксуснокислое.
Молекулярный кислород в процессах брожения не участвует. Большинство бактерий, осуществляющих брожение, — облигатные анаэробы. Однако некоторые из них — факультативные анаэробы — способны осуществлять процесс брожения в присутствии кислорода, но без его участия. Более того, кислород подавляет процесс брожения, и оно сменяется горением (дыханием — конечный акцептор водорода — кислород). Этот эффект был назван эффектом Пастера и является одним из классических примеров смены метаболизма у бактерий в зависимости от условий среды.
3. Синтез биополимеров бактериальной клетки требует энергии. Она образуется в ходе биологического окисления и запасается в в








