 2015-03-20
2015-03-20 5062
5062В энергетическом обеспечении клетки важнейшую роль играет адениловая система, которая включает АМФ, АДФ, Н4Р2О7 (пирофосфат), Н3РО4 (неорганический фосфат) и цАМФ (циклический АМФ). Вопрос об адениловой системе сводится к процессам распада, синтеза АТФ и ее значению для процессов жизнедеятельности клетки. Главным компонентом адениловой системы клетки является АТФ. Это макроэргическое соединение. Как известно, к макроэргическим относятся соединения, при гидролизе которых высвобождается не менее, чем 5 ккал/моль. В ряду макроэргов клетки АТФ отводится главная роль.
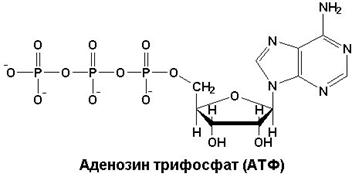
Две фосфоангидридные связи в молекуле АТФ являются макроэргическими. Свободная энергия гидролиза каждой из них равна 7,3 ккал/моль (32 кДж/моль). АТФ обладает высоким потенциалом переноса фосфатных групп на другие вещества (глюкоза, глицерол), тем самым активируя их. С другой стороны, макроэрги, которые имеют больший энергетический потенциал, чем АТФ (например, креатинфосфат), могут переносить свою фосфатную группу на АДФ с образованием АТФ. Таким образом, АТФ занимает центральное положение в ряду других фосфорилированных соединений клетки и является универсальным макроэргом клетки. Это так называемая клеточная энергетическая валюта.
АТФ используется клетками для процессов биосинтеза (анаболические реакции), активации многих молекул (глюкоза, глицерол), выполнения механической работы, переноса веществ через мембраны, обеспечивает точную передачу генетической информации и др. При этом АТФ может гидролизоваться двумя способами:
1) АТФ + Н2О = АДФ + неорганический фосфат + энергия (32 кДж\моль);
2) АТФ + Н2О = АМФ + пирофосфат + энергия (32 кДж\моль).
Синтез АТФ носит название фосфорилирования и описывается уравнением:
АДФ + Н3РО4 = АТФ + Н2О.
Эта реакция происходит при условии обеспечения энергией в количестве не менее
32 кДж/моль.
Если источником этой энергии является транспорт электронов по дыхательной цепи внутренней мембраны митохондрий, говорят об окислительном фосфорилировании. Это главный путь синтеза АТФ в аэробных клетках.
Если источником энергии является гидролиз макроэргической связи субстрата, говорят о субстратном фосфорилировании. Такой механизм имеет место в цитозоле и митохондриях и может происходить в анаэробных условиях.
У растений существует фотосинтетическое фосфорилирование в хлоропластах. Источником энергии в данном случае являются кванты солнечного света.
Следует особо подчеркнуть, что процесс окислительного фосфорилирования тесно связан (сопряжен) с окислительно-восстановительными реакциями (ОВР), а именно с реакцией окисления водорода кислородом до воды — тканевым дыханием. Реакция образования воды in vitro (в пробирке) сопровождается одномоментным выделением около 230 кДж /моль энергии и описывается как реакция взрыва гремучего газа. В живой клетке такой путь термодинамически невозможен, поэтому окисление водорода до воды in vivo (в организме) характеризуется двумя важными особенностями.
Во-первых, газообразный водород в клетках не образуется. Он входит в состав субстратов и отделяется от них путем дегидрирования. Ферменты, которые катализируют эти реакции, — дегидрогеназы (ДГ). Это двухкомпонентные ферменты; они делятся на пиридиновые (ПДГ), которые в качестве кофермента используют производные витамина РР — НАД+ и НАДФ+, и флавиновые, которые в качестве кофермента используют производные витамина В2 — ФМН и ФАД. В ходе ОВР субстраты окисляются, а коферменты восстанавливаются:
Дегидрогеназа
SH2 ¾¾¾¾® S + восстановленный кофермент (НАДН Н+, НАДФН Н+, ФМН·Н2, ФАД Н2)
Кофермент
Во-вторых, выделение энергии происходит постепенно, порциями, для чего процесс окисления водорода осуществляется в несколько стадий с участием ферментов дыхательной цепи. Часть этой энергии запасается в виде АТФ в реакции окислительного фосфорилирования.
Восстановленные субстраты, поставляющие атомы водорода для дыхательной цепи, это небольшие молекулы (карбоновые кислоты, кетокислоты, аминокислоты и др.). Основными поставщиками восстановленных субстратов являются центральные метаболические пути — окислительное декарбоксилирование пировиноградной кислоты и цикл лимонной кислоты. Оба локализованы в матриксе митохондрий, в ходе этих процессов происходят реакции декарбоксилирования (большая часть всей углекислоты, образующейся в клетках, образуется именно здесь). Кроме того, как уже говорилось, в ходе этих процессов происходят реакции дегидрирования субстратов, образуются восстановленные коферментные формы НАДН·Н+ и ФАДН2, водород которых поступает в дыхательную цепь внутренней мембраны митохондрий, где происходит его окисление кислородом до воды и синтез АТФ.
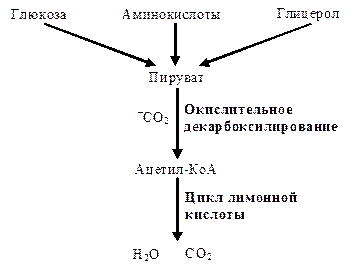
Переходим к рассмотрению центральных метаболических путей.

